|
|
||
 |
 |
 |
|
|
|
Synapse Formation
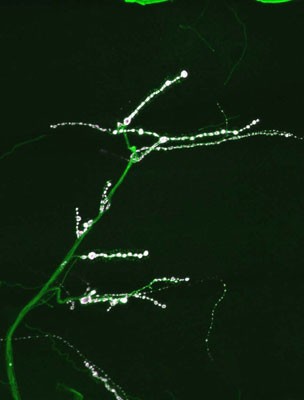
To identify neuronal pathways that control synapse formation and synaptic growth, we have conducted forward genetic screens for temperature-sensitive mutants that disrupt the process. Several Ca2+-dependent presynaptic proteins have been implicated in synaptic growth, but the source of Ca2+ and its regulation are unknown. In addition to retrograde signaling, we have found that presynaptic Ca2+ influx through N-type channels participates in synaptic growth via signaling pathways that are distinct from those that mediate neurotransmitter release. The opening of presynaptic N-type channels during robust synaptic activity allows presynaptic Ca2+ influx to modulate sprouting mechanisms that locally control synaptic remodeling. Linking presynaptic voltage-gated Ca2+ entry to downstream Ca2+-sensitive synaptic growth regulators provides an efficient activity-dependent mechanism for modifying synaptic strength.
Several cell adhesion proteins and synaptic growth regulators reside adjacent to the active zone where presynaptic Ca2+ channels localize, in domains termed peri-active zones. In collaboration with Barry Ganetzky’s lab, we have recently identified one such peri-active zone protein, termed nervous wreck (nwk), in screens for temperature-sensitive paralytic mutants that causes excessive growth of larval neuromuscular junctions. Mutations in nwk result in increased synaptic bouton number and branch formation. NWK encodes an FCH and SH3 domain-containing adaptor protein that localizes to the peri-active zone of presynaptic terminals and binds to the Drosophila ortholog of Wasp (Wsp), a key regulator of actin polymerization. wsp null mutants display synaptic overgrowth similar to nwk and enhance the nwk morphological phenotype in a dose-dependent manner. Evolutionarily, NWK belongs to a previously conserved family of adaptor proteins that includes the human srGAPs, which regulate Rho activity downstream of Robo receptors. We have also found that NWK interacts with the endocytic proteins dynamin and Dap160 and functions together with Cdc42 to promote WASp-mediated actin polymerization in vitro and to regulate synaptic growth in vivo. Cdc42 function is associated with Rab11-dependent recycling endosomes, and Rab11 co-localizes with NWK at the NMJ. Taken together, our results suggest that synaptic growth activated by growth factor signaling is controlled at an endosomal compartment via coordinated NWK and Cdc42-dependent actin assembly.
Further genetic analysis of synaptic growth mutants and NWK interactors is underway to identify additional molecular pathways that modulate synaptic connectivity and alter behavioral output. In addition, genetic screens for mutants disrupting organization of the pre- and post-synaptic region are being conducted to find novel pathways that regulate active zone assembly and pre/post-synaptic alignment. Coupled with new approaches to image synapse formation in live animals in real time, we expect these studies to uncover new molecular pathways that underlie synapse dynamics during development.
|
|