|
|
||
 |
 |
 |
|
|
|
Genetic Dissection of Neurotransmitter Release
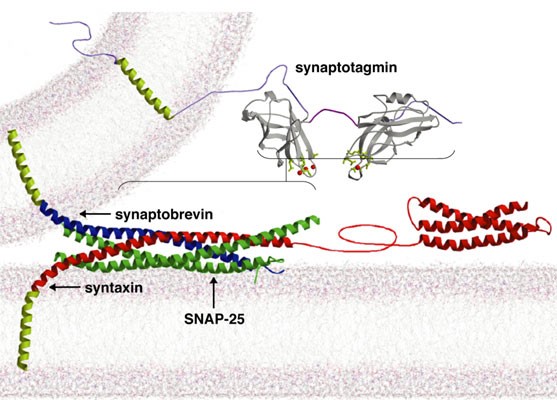
Bernard Katz and colleagues established the hypothesis that Ca2+ influx into the presynaptic nerve terminal triggers neurotransmitter release. Current models for synaptic vesicle exocytosis propose that fusion requires assembly and activation of the SNARE complex. Although SNAREs are essential for synaptic transmission, fusion via reconstituted SNARE proteins occurs over slow timescales and is Ca2+ -independent, contrasting with synaptic transmission, where SNARE assembly and subsequent fusion is rapid and Ca2+ -triggered. A mechanistic understanding of how synaptic Ca2+ signals are transduced into membrane fusion in still lacking. The search for synaptic Ca2+ sensors that regulate SNARE-dependent fusion has largely focused on the synaptotagmins, transmembrane proteins containing tandem Ca2+ binding C2 domains (C2A and C2B). Synaptotagmin1 (Syt 1) is by far the most studied isoform of the synaptotagmin family and contains a small N-terminal intravesicular domain, a single transmembrane domain and a large cytoplasmic region. Two domains homologous to the C2 regulatory domain of protein kinase C occupy the majority of the cytoplasmic region of Syt 1. C2 domains are ~ 130 amino acid motifs that are found in an array of intracellular proteins, many of which bind Ca2+. To characterize the mechanisms that couple Ca2+ influx to rapid and synchronous synaptic vesicle fusion, we are analyzing Drosophila syt 1 mutants deficient in specific interactions mediated by its two Ca2+ -binding C2 domains. In the absence of Syt 1, synchronous release is abolished and a kinetically distinct delayed asynchronous release pathway is uncovered. Mutant synapses containing only the C2A domain of Syt 1 partially recover synchronous fusion, but have an abolished Ca2+ cooperativity. Mutants that disrupt Ca2+ sensing by the C2B domain have synchronous release with normal Ca2+ cooperativity, but with reduced release probability. Our findings indicate Syt 1 is the major Ca2+ sensor for evoked release and functions to trigger synchronous fusion in response to Ca2+, while suppressing asynchronous release. We have also characterized the six remaining synaptotagmin isoforms encoded in the Drosophila genome, including homologs of mammalian Syts 4, 7, 12 and 14. Like Syt 1, Syt 4 is ubiquitously present at synapses but localizes postsynaptically rather than presynaptically. Our results indicate that synaptotagmins are differentially distributed to unique subcellular compartments. Current work is underway to determine the targeting signals that mediate trafficking of the synaptotagmin family to synaptic vesicles, postsynaptic vesicles, or large dense core vesicles. In addition, the function of the remaining synaptotagmin family members is currently being addressed by genetic analysis in Drosophila. Neuronal synapses require rapid fusion of docked synaptic vesicles in response to calcium influx during action potentials. The molecular nature of the fusion clamp machinery that prevents exocytosis of synaptic vesicles in the absence of a calcium signal is still unclear. We have characterized the function of complexin, a small alpha-helical protein that binds fully assembled SNARE complexes, and our data suggests it functions as the synaptic vesicle fusion clamp in vivo. Drosophila has a single complexin homolog that is abundantly expressed in presynaptic nerve terminals. Animals lacking complexin die throughout development, with adult escapers showing severe locomotor defects and a loss of visual function. Electrophysiological analysis at neuromuscular junctions in complexin null mutants reveals a dramatic increase in spontaneous synaptic vesicle fusion that is independent of nerve stimulation or extracellular calcium. The massive increase in spontaneous miniature release is abolished following depletion of synaptic vesicles in shibire, complexin double mutants. These data indicate that complexin interacts with assembled SNARE complexes to prevent premature vesicle fusion in the absence of calcium entry. Future work is aimed at understanding how complexin regulates both spontaneous and evoked release, and the role of spontaneous mini release in synaptic biology.
|
|