![]()
--> Home Page | Report Series |
X. Xiao 1,2, J.M. Melillo 1, D.W. Kicklighter 1, Y. Pan 1, A.D. McGuire 3 and J. Helfrich 1
October 1996
View Report as PDF file (65 kB)
Contents
Atmospheric concentrations of CO2 and other long-lived greenhouse gases (e.g., N2O) will continue to increase in the next century as the result of increasing anthropogenic emissions of these trace gases. Increasing greenhouse gases will further increase the radiative forcing of climate. For doubled CO2, a number of atmospheric general circulation models (GCMs) estimate that globally averaged surface air temperature will increase in the range of 1.5 degrees C to 4.5 degrees C (Mitchell, et al., 1990). Globally, there are also large geographical variations in the changes of precipitation and clouds projected by GCMs. The increasing atmospheric CO2 concentration and resultant climate change are likely to have significant impacts on net primary production of terrestrial ecosystems (Melillo, et al., 1990). Net primary production of terrestrial ecosystems is important in estimating land carrying capacity, which is critically relevant to the social and economic development of a country like China. China has a very large human population (about 1/5 of the world population) and limited natural resources, especially fertile lands. The total land area in China is only about 1/15 of the world land area and about 65% of this area is hills, mountains and plateau (Xiong and Li, 1988).
In China, the magnitude and spatial patterns of temperature and precipitation will change significantly according to simulations of a GCM for a doubling of atmospheric CO2 concentration (Zhang and Wang, 1993; Wang and Zhang, 1993). Few studies have examined impacts of climate change on net primary production (NPP) of natural ecosystems in China (Zhang, 1993; Xiao, et al., 1995). Zhang (1993) applied a regression model (Chikugo model, Uchijima and Seino, 1985), which uses annual values of climate variables (e.g., temperature, precipitation, relative humidity) to estimate NPP of various ecosystems in China for contemporary climate. Zhang (1993) also calculated the responses of NPP to: (1) +2 degrees C and 20% increase of annual precipitation, and (2) +4 degrees C and 20% increase of annual precipitation in China. Net primary production increases for all the vegetation types under these climate change conditions (Zhang, 1993). However, the Chikugo model is basically a regression model based on correlation between NPP and climate, and the model has not considered the possible limitations of nutrient availability (e.g., nitrogen) on NPP. Generally, the potential of application of regression models for future projection is limited because the regressions may not be appropriate for climatic conditions that are novel to terrestrial ecosystems (Melillo, et al., 1993).
Recently, a number of processed-based ecosystem models, which are integrated with geographically referenced spatial data, have been applied to estimate responses of NPP and carbon storage to changes in climate and atmospheric CO2 concentration at the global scale (Melillo, et al., 1993; McGuire, et al., 1995, 1996; Xiao, et al., 1996a) and the continental scale, e.g., South America (Raich, et al., 1991), North America (McGuire, et al., 1992, 1993) and conterminous United States (VEMAP Members, 1995). Estimates of NPP responses vary among the GCMs used in the studies across the scales of grid cell, biome, continent and the globe (Melillo, et al., 1993; VEMAP, 1995; Xiao, et al., 1996a). The process-based ecosystem models integrate key ecosystem processes such as plant photosynthesis, plant respiration, decomposition of soil organic matter and nutrient cycling, which interactively affect NPP. These ecological processes are controlled by a number of abiotic factors, e.g., water, light, temperature, soil texture and nutrients. The simultaneous interactions among the dynamics of carbon, nitrogen and water are complex and spatially variable.
In this study, we used a process-based global terrestrial biogeochemistry model, the Terrestrial Ecosystem Model (Raich, et al., 1991; McGuire, et al., 1995, 1996). First, we applied TEM to estimate the magnitude and spatial distribution of annual NPP of terrestrial ecosystems in China for contemporary climate. Second, we applied TEM to quantify the equilibrium responses of annual NPP in China to changes in atmospheric CO2 concentration and climate projected by three GCMs. We examined the TEM simulation results across the spatial scales of grid cell, biome and continent. The analysis will help us to understand how the abiotic factors (e.g., climate, soil texture) control the magnitude and spatial distribution of NPP in China.
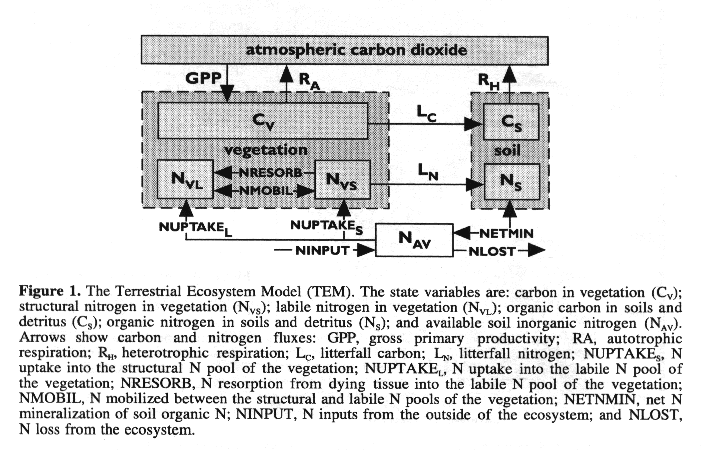
The TEM (Figure 1) is a process-based ecosystem model that simulates important carbon and nitrogen fluxes and pools for various terrestrial biomes (Raich, et al., 1991; McGuire, et al., 1992, 1993, 1995). The TEM has been used to examine patterns of net primary production (NPP) in South America (Raich, et al., 1991) and North America (McGuire, et al., 1992) for contemporary climate. The model has also been used to estimate the responses of NPP of the terrestrial biosphere to climate change projected by four GCMs (Melillo, et al., 1993). In this study, we use version 4.0 of TEM, which was modified from TEM version 3 to improve patterns of soil organic carbon storage along gradients of temperature, moisture and soil texture (McGuire, et al., 1995, 1996). Version 4.0 of TEM has also been used to estimate the responses of NPP and carbon storage of terrestrial ecosystems to climate change in the conterminous United State (VEMAP Members, 1995; Pan, et al., 1996a) and at the global scale (McGuire, et al., 1996; Xiao, et al., 1996a, 1996b). Detailed descriptions of TEM are documented in Raich, et al. (1991) and McGuire, et al. (1992). The recent TEM modifications are presented elsewhere (McGuire, et al., 1995, 1996). To help understand how climate change and elevated CO2 influence NPP in TEM, we review the key processes related to NPP.
Net primary production (NPP) is calculated as
the difference between gross primary production (GPP) and plant
respiration (R). The flux GPP is calculated at each
monthly time step as follows: GPP = C
The driving variables for the spatial extrapolation of TEM are vegetation types, elevation, soil texture, monthly mean temperature, monthly precipitation, monthly mean cloudiness, and several hydrological variables (potential evapotranspiration, estimated evapotranspiration and soil moisture). Hydrological inputs for TEM are generated by a water balance model (Vorosmarty, et al., 1989) that uses the same climate, elevation, soils and vegetation data.
In this study, the spatial data used to drive TEM are derived from our global data sets (Melillo, et al., 1993; McGuire, et al., 1995, 1996; Xiao, et al., 1996a), which are organized in a grid at a 0.5 degrees (latitude) x 0.5 degrees (longitude) resolution. China has 3,852 grid cells, including six grid cells of inland water body and 26 grid cells of wetlands. The TEM does not estimate NPP and carbon storage for grid cells defined as water or wetland ecosystems, so the extrapolation of TEM to China requires the application of TEM to 3820 grid cells.
We use potential vegetation in this study. Our global classification of potential vegetation includes 18 upland biomes (Melillo, et al., 1993). In developing the global vegetation data, the Vegetation Map of China (Hou, et al., 1982) was used in digitization and aggregation. There are 16 upland biomes in China (Plate 1). Forests dominate in southern China and eastern coastal areas, while arid and semi-arid biomes prevail in northwestern China. The Qinhai-Tibet plateau is dominated by polar desert/alpine tundra.
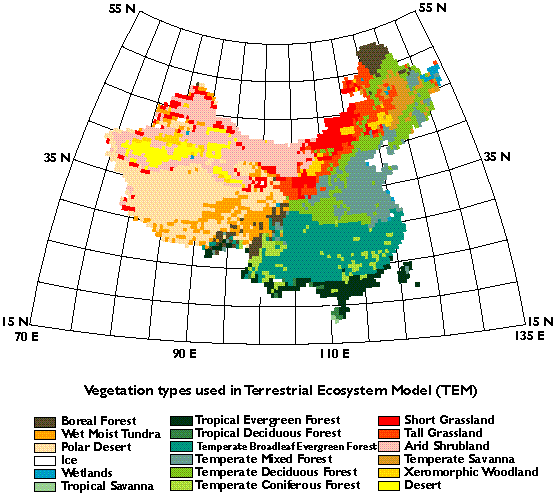
Plate 1. Geographical
distributions of potential natural vegetation in China.
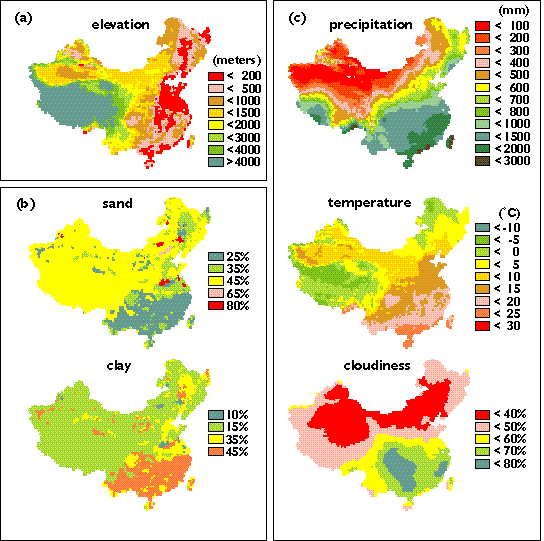
Table 1. Proportions of sand, silt and clay assigned to the FAO/CSRC/MBL soil texture class.
We first ran TEM for contemporary climate at 312.5 ppmv CO2 as the baseline for comparison. We used 312.5 ppmv CO2 because it is the average value of atmospheric CO2 concentration for the 1 x CO2 simulations used by the GCMs described in Melillo, et al., (1993). To help separate the relative importance of elevated CO2 and climate changes, we then ran TEM under (1) elevated CO2 only, (2) climate change only, and (3) climate change plus elevated CO2. For each of the four simulations, we ran TEM to steady state, therefore, its estimates of annual NPP apply only to mature, undisturbed vegetation and ecosystems. We have not considered the impacts of changes in land use and land cover on NPP in this study.
For climate change predictions, we used climate outputs for "current" (1 x CO2) and "doubled" (2 x CO2) simulations from three GCMs, i.e., Geophysical Fluid Dynamic Laboratory (GFDL-q, Weatherald and Manabe, 1988), Goddard Institute for Space Studies (GISS, Hansen, et al., 1983, 1984) and Oregon State University (OSU, Schlesinger and Zhao, 1989). These GCMs have very coarse spatial resolutions, i.e., 10.0 degrees (longitude) x 7.83 degrees (latitude) for GISS, 7.5 degrees x 4.44 degrees for GFDL and 5.0 degrees x 4.00 degrees for OSU. In order to be consistent with the spatial resolution of the contemporary climate data, the outputs of the GCMs were interpolated to a 0.5 degree x 0.5 degree grid resolution by applying a spherical interpolation routine (Willmott, et al., 1985) to the data.
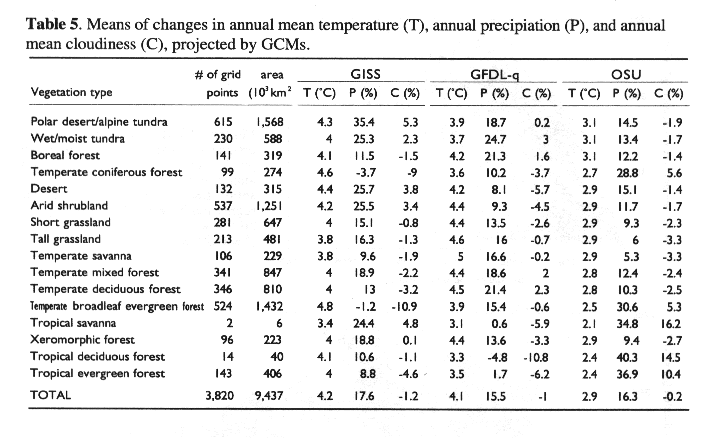
We calculated the differences in monthly mean temperature and the ratios in monthly precipitation and mean monthly cloudiness between the 2 x CO2 and 1 x CO2 simulations of the GCMs. The OSU climate has the smallest changes in annual mean temperature and annual precipitation, while the GISS climate has the largest changes of annual mean temperature, annual precipitation and annual mean cloudiness. The projected climate changes are the most uniform across China in the OSU climate, whereas the GISS climate has large spatial variations in the magnitude of projected climate changes (Plate 3). Spatial distributions of projected changes in annual mean temperature, annual precipitation and annual mean cloudiness vary among the GCMs (Plate 3).
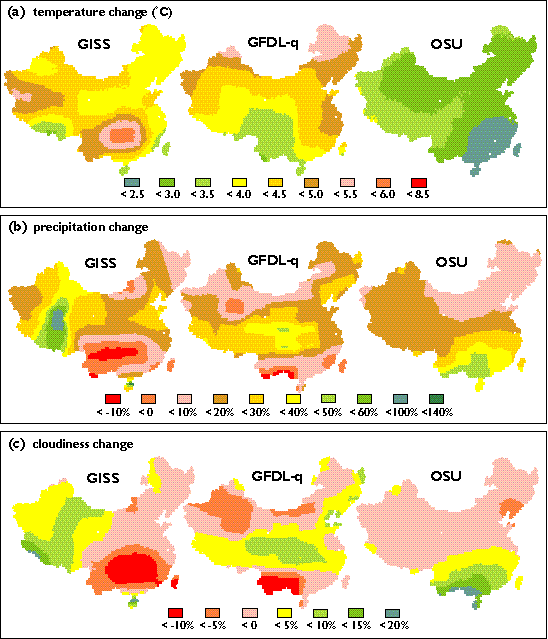
For climate change with 312.5 ppmv CO2, TEM estimates that the continental response of annual NPP in China ranges from a decrease of 54.8 TgC yr-1 (-1.5%) for the GISS climate to an increase of 306.9 TgC yr-1 (8.4%) for the GFDL-q climate (Table 4). These differences in NPP response are determined by the differences among the three GCM climates. According to the GCMs projections, changes in annual mean temperature in China ranges from an increase of 2.9 degrees C for the OSU climate to an increase of 4.2 degrees C for the GISS climate (Table 5). Annual precipitation increases considerably but varies little among the GCMs, i.e. 15.5% for the GFDL-q climate to 17.6% for the GISS climate (Table 5). However, annual mean cloudiness declines slightly, ranging from -0.2% for the OSU climate to -1.2% for the GFDL-q climate (Table 5).
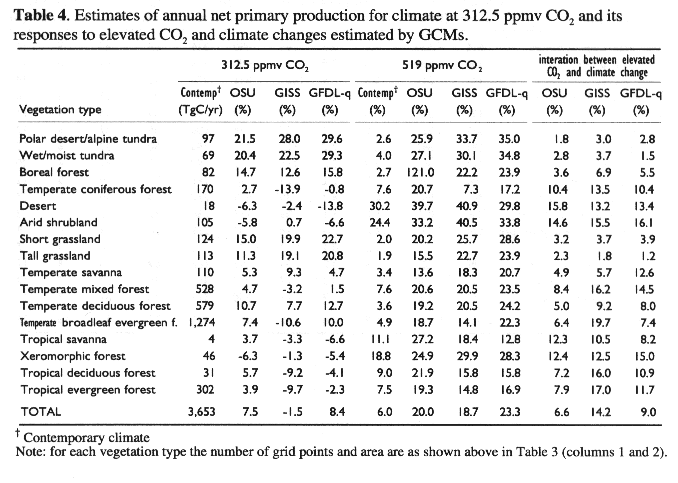
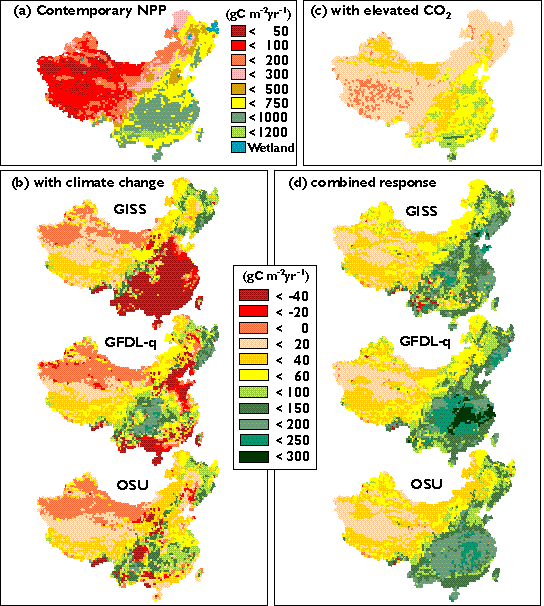
For contemporary climate with elevated CO2 (519 ppmv), TEM estimated that annual NPP increases by 219 TgC yr-1 (6.0%) in China. In TEM, a direct effect of elevated atmospheric CO2 level is to increase the intercellular CO2 concentration within the canopy, which potentially increases GPP via a Michaelis-Menton (hyperbolic) function. Elevated atmospheric CO2 level may also indirectly affect GPP by altering the carbon-nitrogen status of the vegetation increase effort toward nitrogen uptake; increased effort is generally realized only when GPP is limited more by carbon availability than by nitrogen availability (VEMAP Members, 1995).
At the biome scale, annual NPP increases for all of the 16 biomes (Table 4). The NPP responses vary substantially among biomes, ranging from an increase of 1.9% in tall grasslands to an increase of 30.2% in desert (Table 4). Biomes in the arid regions (desert, arid shrubland) have the largest percentage increase in annual NPP. This is attributed mostly to a significant increase of water use efficiency of plants in the arid regions, where water is the primary limiting factor on NPP. Temperate forests and boreal forests are generally limited by the availability of inorganic nitrogen in the soils, and as the result, their responses to elevated CO2 is strongly constrained by nitrogen availability. Tropical savanna, xeromorphic forest, tropical deciduous forest and tropical evergreen forest have large increases of annual NPP in the range of 7.5% to 18.8%. In the tropical regions, annual NPP of ecosystems is generally not limited by nitrogen, because of high rates of net N mineralization resulting from high temperatures.
The geographical distribution of NPP responses to elevated CO2 has distinct spatial patterns (Plate 4c). Annual NPP increase from about 20 gC m^-2yr-1 to 300 gC m^-2yr-1 in Southeastern China. Annual NPP in arid regions increases in the range of 20 to 40 gC m^-2yr-1. In the Qinhai-Tibet Plateau, the response of annual NPP is within ±20 gC m^-2yr-1 (Plate 4c). Annual NPP in the Qinhai-Tibet Plateau is primarily controlled by low temperature and constrained by low nutrient availability.
At the continental scale, annual NPP of China for climate change at 519 ppmv CO2 increases substantially but varies little among the GCM climates, ranging from 683 TgC yr-1 (18.7%) for the GISS climate to 851 TgC yr-1 (23.3%) for the GFDL-q climate. Annual NPP increases considerably for all of the 16 biomes among the three GCM climates (Table 4). Dryland ecosystems (desert, arid shrubland) have the largest percentage increases in annual NPP (more than 30%). Annual NPP in tundra and boreal forest also increases substantially (more than 20%). Although the percentage increases in annual NPP for temperate broadleaf evergreen forest are moderate, this biome accounts for the largest portion of the increase of annual NPP in China: 238.2 TgC yr-1 for the OSU climate, 179.6 TgC yr-1 for the GISS climate and 284.1 TgC yr-1 for the GFDL-q climate.
Geographically, the responses of annual NPP decrease from southeast to northwest within a GCM climate (Plate 4d). The spatial patterns of NPP responses are generally similar among the three GCM climates. Annual NPP has a slight increase (<40 gC m^-2yr-1) in the Qinhai-Tibet plateau and arid regions, a moderate increase (40 to 60 gC m^-2yr-1) in grasslands, and a large increase (> 60 gC m^-2yr-1) in forest regions (Plate 4d). Increases of annual NPP in forest regions are the largest in the GFDL-q climate, up to 200-300 gC m^-2yr-1 in the southern China. For the GISS climate, NPP decreases in scattered areas of the Southern China (Plate 4). There are slight differences in both the magnitude and spatial distributions of NPP responses in the transitional zones between forests and grasslands among the three GCM climates (Plate 4).
It is important to note that at the continental scale, the response of NPP to climate change at 519 ppmv CO2 is much larger than the sum of the NPP response to elevated CO2 only and the NPP response to climate change only (Table 4). For the OSU climate, the sum of the NPP response for climate change at 315 ppmv CO2 (7.5%) and the NPP response for contemporary climate at 519 ppmv CO2 (6.0%) is much lower than the 20.0% NPP response for climate change at 519 ppmv CO2; the difference is attributed to the interaction between climate change and elevated CO2. Similarly, the interaction between climate change and elevated CO2 contributes about 9.0% to the overall response of NPP to climate change at 519 ppmv CO2 for the GFDL-q climate and 14.2% for the GISS climate. The interaction between elevated CO2 and climate change is primarily caused by enhanced plant nitrogen uptake and increasing availability of mineralized nitrogen and carbon (CO2) resources. These results show clearly that the interaction between elevated CO2 and climate change contributes significantly to the overall equilibrium response of NPP to changes in climate and CO2 (Table 4).
The relative role of the interaction between elevated CO2 and climate change varies among the 16 biomes, ranging from a large role in arid biomes (e.g., desert, arid shrubland) to a small role in alpine tundra (Table 4). The relative role of the interaction between elevated CO2 and climate change also varies among the three GCMs climates (Table 4). This indicates that geographical distributions of climate changes projected by the GCMs affect the interaction between elevated CO2 and climate change at larger spatial scales.
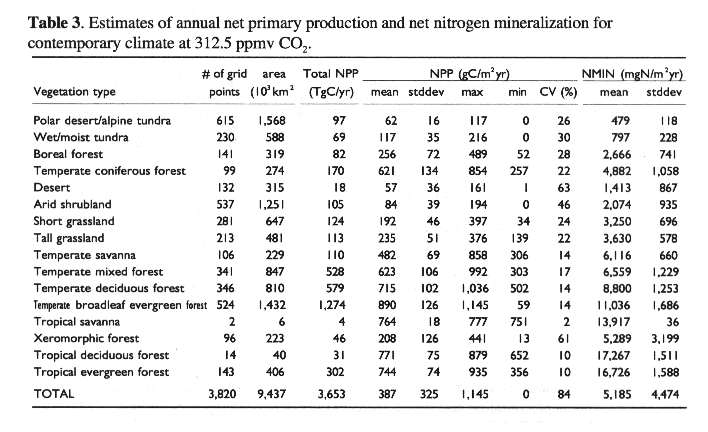
There are numerous field measurements of plant biomass and NPP for various ecosystems in China, however, few studies have estimated the spatial distribution of NPP in China on large spatial scales. Zhang (1993) calculated annual NPP for 671 weather stations in China, using the Chikugo model (Uchijima and Seino, 1985). Fang, et al., (1995) and Xiao, et al., (1996) estimated annual NPP in China in 1990, using Normalized Difference Vegetation Index (NDVI) derived from remote sensing data of NOAA AVHRR. In our study, the TEM-estimated annual NPP of all the terrestrial ecosystems in China under contemporary climate at 312.5 ppmv CO2 is 3,653 TgC yr-1, about 8% of global terrestrial NPP (see Melillo, et al., 1993). Application of process-based ecosystem models such as TEM provides an important and useful tool to study spatial patterns and dynamics of NPP in China, although NPP estimates can be improved with the collection of additional field data and remote sensing data to further develop and validate ecosystem models.
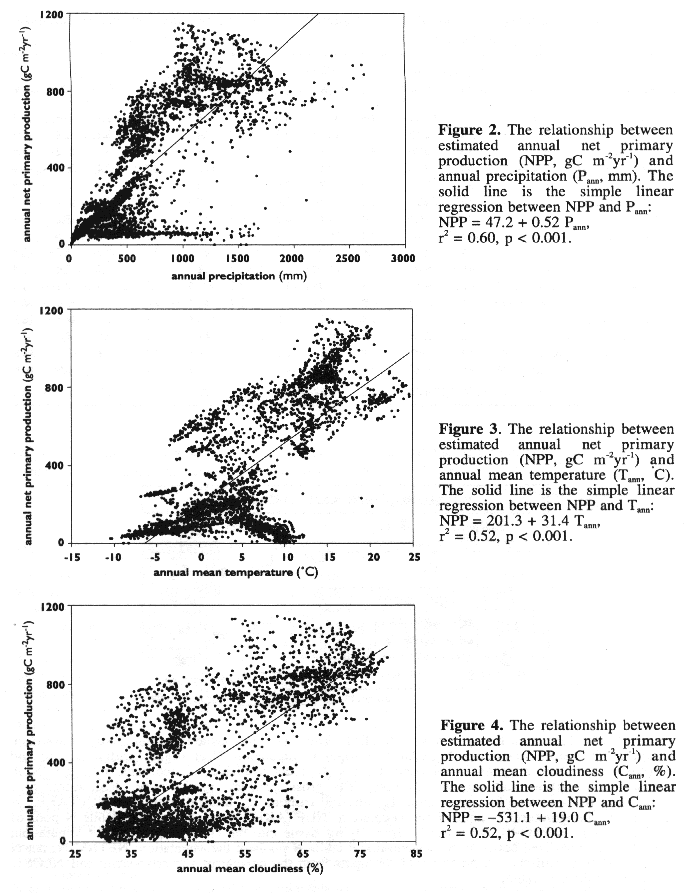
The TEM results show that annual NPP in China has a large spatial variation. The geographical distribution of annual NPP in this study is similar to Zhang (1993) and Xiao, et al. (1996), characterized by a gradient of high NPP in southeastern China to low NPP in northwestern China. The spatial patterns of NPP are closely correlated to the spatial distributions of abiotic factors, particularly climate. This indicates that the spatial representation of contemporary climate and other abiotic factors used in extrapolation of TEM for China may affect estimates of NPP. In an earlier study that examines three alternative geo-referenced data sets of climate, solar radiation and soil texture in estimating NPP of the conterminous United States, Pan, et al. (1996a) found that NPP estimates of TEM are sensitive to variations in the input data, and the relative importance of climate, soil texture and solar radiation to NPP estimates differs among the vegetation types.
The spatial input data used in this study are from our global data base and need to be updated with the new data available in China. For instance, the climate data used in this study are the long-term average values of weather records from about 300 weather stations in China (W. Cramer, personal communication). Leemans and Cramer (1990) compiled weather record data mainly for the period of 1931-1960. Zhang (1993) calculated biotemperature and potential evapotranspiration using long-term average data from about 671 weather stations in China. Using more weather stations in the spatial interpolation will improve the spatial and temporal (seasonal) representations of contemporary climate.
Major soils in Xinjian, Qinhai-Tibet plateau and Inner Mongolia have generally about 40 to 70% of sand proportion in soil texture (Xiong and Li, 1987), which is higher than the 35 or 45% of sand proportion we used in this study to drive TEM (Plate 2). In arid and semi-arid regions, coarse texture soils, which allow deep infiltration of water, may have lower evaporation but higher transpiration rate than fine texture soils (Noy-Meir, 1973; Sala, et al., 1988). Primary production is therefore higher in coarse texture soils than in fine texture soils in arid and semi-arid regions (Noy-Meir, 1973). In grasslands of the Central Plains of United States, aboveground production increases with coarser soil texture when annual precipitation is less than 370 mm, but decreases when annual precipitation is greater than 370 mm (Sala, et al., 1988).
Therefore, to improve estimates of NPP in China, geographically referenced spatial data with better spatial-temporal representations are critically needed. Improved data set will allow ecosystem models such as TEM to make more realistic estimates of NPP for contemporary climate. As the contemporary climate data sets are also used in generating a "future climate" for impact analyses of climate change, improved contemporary climate data sets are likely to reduce the uncertainty in estimating responses of NPP in China to climate change.
The TEM results show that NPP of terrestrial ecosystems in China is sensitive to changes in atmospheric CO2 concentration and climate as projected by the GCMs. These results are consistent with an earlier study (Xiao, et al., 1995) that used the CENTURY ecosystem model (Parton, et al., 1993) at individual sites and showed that NPP in grasslands of Inner Mongolia of China is sensitive to changes in climate and atmospheric CO2 concentration. The TEM results also show that the magnitude and spatial distribution of NPP responses vary considerably among the GCMs projections. The largest uncertainty in the NPP response to climate change among the three GCM climates occurs in Southern China. The uncertainty in NPP responses is related to the uncertainty in the GCMs projections for future climate change.
There is a large uncertainty in the GCMs projections for future climate changes. A number of GCMs are available (Mitchell, et al., 1990). Most GCMs project a similar range of global surface temperature increase, but they vary significantly in the spatial patterns of projected changes in temperature and precipitation. Zhang and Wang (1993) examined the simulation results for 1 x CO2 and 2 x CO2 scenarios from the Community Climate Model (CCM) at the National Center for Atmospheric Research (NCAR) of the United States. Annual mean temperature in China increases by 2.5-3.0 degrees C. There is a large spatial variation in changes of annual precipitation in China. In particular, the NCAR CCM predicts a considerable decrease (about 200 mm) of annual precipitation in the semi-arid Loess Plateau, where serious soil-water erosion and land degradation occur extensively.
Because of the very coarse spatial resolutions of GCMs, GCMs are generally poor in representing regional climate change, especially in China, which has very complex topography and the climate is dominated by summer monsoon. Monsoon climate is characterized by seasonal jumps, large interannual variability and abrupt changes on decade and longer time scales. There are strong interactions between climate and ecosystems in the Asian monsoon region on various time scales (Fu, 1995a). For instance, the normalized difference vegetation index (NDVI) for southern China varied significantly in the period of 1985-1990 (Fu, 1995a). The coupling of GCMs and meso-scale climate models with detailed representations of topography and ecosystem dynamics may generate better spatial-temporal representations of climate change in China (Fu, 1995b).
In this study, we have not considered the responses of vegetation distribution to climate change and elevated CO2. Climate change is likely to affect the spatial distribution of vegetation in China (Zhang and Liu, 1993; Zhang and Song, 1993; Zhang, 1993; Wang and Zhao, 1995; Zhou and Zhang, 1996). Vegetation redistribution can affect regional estimates of NPP of terrestrial ecosystems (VEMAP Members, 1995). How climate change and elevated CO2 affect vegetation distribution in China and consequently NPP is discussed in another paper (Pan, et al., 1996b).
We used the potential vegetation of China in this study. Thus, we have not considered the effects of land use and management on NPP in China. In the last several thousand years of agricultural practice, a large portion of uplands has been converted to croplands. According to the Vegetation Map of China (Hou, et al., 1982), the area of croplands is approximately 1.5 million km^2, or about 15% of the total land area of China. Based on the Land Use Map of China at the scale of 1:1,000,000 (Wu, 1990), cultivated lands account for about 14.2% of the total land area of China (Wu and Guo, 1994). Conversion of forests to croplands results in significant losses of NPP and carbon storage (Houghton and Skole, 1990). Climate change is likely to have significant impacts on physical geographical zones (Zhao, 1993) and agriculture in China (Wang, 1993; Gong and Hameed, 1993; Ohta, et al., 1995). This study provides a baseline for future studies to investigate the impacts of changes in land use and land cover on NPP and carbon storage of China.
In summary, this study demonstrates the usefulness of integrating a process-based ecosystem model with geo-referenced spatial data in China for examining the spatial patterns and dynamics of net primary production of terrestrial ecosystems. However, the analyses in this study are based on an equilibrium changes in climate and carbon fluxes of terrestrial ecosystems. The time course of changes in atmospheric CO2 concentration, climate, land use and anthropogenic nitrogen deposition from the last several centuries to next century have not been considered. We are beginning to conduct transient simulations of TEM to track the path and magnitude of the responses of NPP over time (Melillo, et al., 1996), but progress is hindered by a lack of both historical and future data with better spatial-temporal representation on climate, land use and N deposition in China. As these data sets become available in the near future, we should be able to improve the estimates of NPP and to better assess the effects of land use and climate changes on NPP of terrestrial ecosystems in China.
The study was supported by the National Aeronautics and Space Administration's Earth Observing System (NAGW-2669), the Department of Energy's National Institute for Global Environmental Change (No: 901214-HAR) and the Joint Program on Science and Policy of Global Change at the Massachusetts Institute of Technology (CE-S-462041).
Fang, Z., Q. Xiao, Y. Liu and R. Sheng, 1995,
Monitoring of some surface properties in China by NOAA AVHRR.
In: "China Contributions to Global Change
Studies," D. Yu and H. Lin (eds.), Science Press, Beijing,
p. 146-151.
FAO/CSRC/MBL (undated), Soil Map of the World,
1:5,000,000, Unesco, Paris, France. Digitization (0.5 degree
resolution) by Complex Systems Research Center, University of New
Hampshire, Durham, and modifications by Marine Biological
Laboratory, Woods Hole, MA.
FAO/UNESCO, 1971, Soil Map of the World, 1:5,000,000,
Unesco, Paris, France.
Fu, C., 1995a, Dynamics of monsoon-driven ecosystem:
Concept, preliminary evidence and proposal for further research. In:
"China Contributions to Global Change Studies," D. Yu
and H. Lin (eds.), Science Press, Beijing, p. 103-106.
Fu, C., 1995b, Simulation of summer monsoon in east China
by high resolution regional climate-ecosystem model (RCEM), In:
"China Contributions to Global Change Studies," D. Yu
and H. Lin (eds.), Science Press, Beijing, p. 142-145.
Gates, D.M., 1985, Global biospheric response to
increasing atmospheric carbon dioxide concentration. In:
"Direct Effect of Increasing Carbon Dioxide on
Vegetation," B.R. Strain and J.D. Cure (eds.), DOE/ER-0238,
United States Department of Energy, Washington D.C., p. 171-184.
Gong, G., and S. Hameed, 1993, Identification of
climatically sensitive agricultural zones of China. In:
"Climate Change and Its Impact," Y. Zhang, P. Zhang, H.
Zhang and Z. Lin (eds.), Meteorology Press, Beijing, p. 78-90.
(in Chinese with English abstract)
Hansen, J., G. Russel, D. Rind, P. Stone, A. Lacis, S.
Lebedeff, R. Ruedy and L. Travis, 1983, Efficient three
dimensional global models for climate studies: Model I and II, Monthly
Weather Review, 111:609-662.
Hansen, J., A. Lacis, D. Rind, G. Russel, P. Stone, I.
Fung, R. Ruedy and J. Lerner, 1984, Climate sensitivity: Analysis
of feedback mechanisms. In: "Climate process and
Climate Sensitivity," J.E. Hansen and T. Takahashi (eds.),
Geophysical Monograph 29, Maurice Ewing series 5, American
Geophysical Union, Washington, D.C., p. 130-163.
Hou, X. (ed.), 1982, Vegetation map of China, 2nd edition,
Science Press, Beijing. (in Chinese)
Houghton, R.A., and D.L. Skole, 1990, Changes in the
global carbon cycle between 1700 and 1985. In: "The
Earth as Transformed by Human Action," B.L. Turner, et al.
(eds.), Cambridge University Press, Cambridge, UK, p. 393-408.
IPCC (Intergovernmental Panel on Climate Change), 1995, Climate
Change 1994: Radiative forcing of climate change and an
evaluation of the IPCC IS92 emission scenarios, Cambridge
University Press, New York, p. 196-197.
Jensen, M.E., and H.E. Haise, 1963, Estimating
evapotranspiration from solar radiation, Journal of the
Irrigation and Drainage Division, 4:15-41.
Kimball, B.A., 1975, Carbon dioxide and agricultural
yield: An assemblage and analysis of 430 prior observations, Agronomy
Journal, 75:779-788.
Leemans, R., and W. P. Cramer, 1990, The IIASA climate
database for land areas on a grid with 0.5 degree resolution,
WP-90-41, International Institute for Applied Systems Analysis,
Laxenburg, Austria, 60 pp.
Manabe, S., and R.T. Wetherald, 1987, Large scale changes
in soil wetness induced by an increase in carbon dioxide, Journal
of the Atmospheric Sciences, 44:1211-1235.
McGuire, A.D., J.M. Melillo, D.W. Kicklighter, Y. Pan, X.
Xiao, J. Helfrich, B. Moore, III, C.J. Vorosmarty and A.L.
Schloss, 1996, The role of the nitrogen cycle in the global
response of net primary production and carbon storage to doubled
atmospheric carbon dioxide, Global Biogeochemical Cycles
(in review).
McGuire, A.D., J.M. Melillo, D.W. Kicklighter and L.A.
Joyce, 1995, Equilibrium response of soil organic carbon to
climate change: empirical and process-based estimates, Journal
of Biogeography, 22:785-796.
McGuire, A.D., L.A. Joyce, D.W. Kicklighter, J.M. Melillo,
G. Esser and C.J. Vorosmarty, 1993, Productivity response of
climax temperate forests to elevated temperature and carbon
dioxide: A North America comparison between two global models, Climatic
Change, 24:287-310.
McGuire, A.D., J.M. Melillo, L.A. Joyce, D.W. Kicklighter,
A.L. Grace, B. Moore, III and C.J. Vorosmarty, 1992, Interactions
between carbon and nitrogen dynamics in estimating net primary
productivity for potential vegetation in North America, Global
Biogeochemical Cycles, 6(2):101-124.
Melillo, J.M., I.C. Prentice, G.D. Farquhar, E.D. Schulze
and O.E.Sala, 1996, Terrestrial biotic responses to environmental
change and feedback to climate. In: "IPCC Climate
Change 1995: The Science of Climate Change,"
Intergovernmental Panel on Climate Change, Cambridge University
Press, p. 445-481.
Melillo, J.M., 1994, Modeling land-atmospheric
interaction: A short review. In: "Changes in Land
Use and Land Cover: A Global Perspective," W.B. Meyer and
B.L. Turner (eds.), Cambridge University Press, p. 387-409.
Melillo, J.M., A.D. McGuire, D.W. Kicklighter, B. Moore,
III, C.J. Vorosmarty and A.L. Schloss, 1993, Global climate
change and terrestrial net primary production, Nature, 363:234-240.
Mitchell, J.F.B., S. Manabe, V. Meleshko and T. Tokioka,
1990, Equilibrium climate change and its implications for the
future. In: "Climate Change: The IPCC Scientific
Assessment," J.T. Houghton, G.J. Jenkins and J.J. Ephraums
(eds.), Cambridge University Press, New York, p. 131-172.
NCAR/Navy, 1984, Global 10-minute elevation data. Digital
tape available through National Oceanic and Atmospheric
Administration, National Geophysical Data Center, Boulder, CO.
Noy-Meir, I., 1973, Desert ecosystems: Environment and
producers, Annual Review of Ecology and Systematic, 4:25-51.
Ohta, S., Z. Uchijima and W. Oshima, 1995, Effect of 2 x
CO2 climatic warming on water temperature and agricultural
potential in China, Journal of Biogeography, 22:649-655.
Pan, Y., A.D. McGuire, D.W. Kicklighter and J.M. Melillo,
1996a, The importance of climate and soils on estimates of net
primary production: A sensitivity analysis with the Terrestrial
Ecosystem Model, Global Change Biology, 2:5-23.
Pan, Y., J.M. Melillo, D.W. Kicklighter, X. Xiao and A.D.
McGuire, 1996b, Potential response of net primary production in
terrestrial ecosystems of China to climate change: A simulation
study by Terrestrial Ecosystem Model coupled with vegetation
redistribution, Journal of Biogeography ( in
preparation).
Parton, W.J., J.M.O. Scurlock, D.S. Ojima, T.G. Gilmanov,
R.J. Scholes, Schimel, T. Kirchner, J.C. Menaut, T. Seastedt, T.
Garcia Moya, A. Kamnalrut and J.I. Kinyamario, 1993, Observation
and modeling of biomass and soil organic matter dynamics for the
grassland biome worldwide, Global Biogeochemical Cycles,
7(4):785-809.
Prinn, R., H. Jacoby, A. Sokolov, C. Wang, X. Xiao, Z.
Yang, R. Eckaus, P. Stone, D. Ellerman, J. Melillo, J.
Fitzmaurice, D. Kicklighter, Y. Liu and G. Holian, 1996,
Integrated global system model for climate policy analysis: I.
Model framework and sensitivity studies, Joint Program on the
Science and Policy of Global Change, Report No. 7,
Massachusetts Institute of Technology, Cambridge, MA, 76 pp.
Raich, J., W., E.B. Rastetter, J.M. Melillo, D.W.
Kicklighter, P.A. Steudler, B.J. Peterson, A.L. Grace, B. Moore,
III and C.J. Vorosmarty, 1991, Potential net primary productivity
in south America: Application of a global model, Ecological
Applications, 1(4):399-429.
Sala, O.E., W.J. Parton, L.A. Joyce and W.K. Lauenroth,
1988, Primary production of the central grasslands region of the
United States, Ecology, 69:40-45.
Uchijima, Z., and H. Seino, 1985, Agroclimatic evaluation
of net primary productivity of natural vegetations: (1) Chikugo
model for evaluating net primary productivity, Journal of
Agricultural Meteorology, 40(4):343-352.
VEMAP Members, 1995, Vegetation/Ecosystem modeling and
analysis project: Comparing biogeography and biogeochemistry
models in a continental-scale study of terrestrial ecosystem
responses to climate change and CO2 doubling, Global
Biogeochemical Cycles, 9(4):407-437.
Vorosmarty, C.J., B. Moore, III, A.L. Grace, M.P. Gildea,
J.M. Melillo, B.J. Peterson, E.B. Rastetter and P.A. Steudler,
1989, Continental scale models of water balance and fluvial
transport: an application to South America, Global
Biogeochemical Cycles, 3(3):241-265.
Wang, B., 1993, How to deal with the problem of global
warming. In: "Climate Change and Its Impact,"
Y. Zhang, P. Zhang, H. Zhang and Z. Lin (eds.), Meteorology
Press, Beijing, p. 1-15. (in Chinese with English abstract)
Wang, F., and Z. Zhao, 1995, Impact of climate change on
natural vegetation in China and its implication for agriculture, Journal
of Biogeography, 22:657-664.
Wang, W., and Y. Zhang, 1993, The potential change of
precipitation of China under the condition of globe warming
induced by CO2 doubling. In: "Climate Change and
Its Impact," Y. Zhang, P. Zhang, H. Zhang and Z. Lin (eds.),
Meteorology Press, Beijing, p. 257-265. (in Chinese with English
abstract)
Wetherald, R.T., and S. Manabe, 1988, Cloud feedback
processes in a general circulation model, Journal of the
Atmospheric Sciences, 45:1397-1415.
Willmott, C.J., M.R. Clinton and W.D. Philpot, 1985,
Small-scale climate maps: A sensitivity analysis of some common
assumptions associated with grid-point interpolation and
contouring, The American Cartographer, 12:5-16.
Wilson, C.A., and J.F.B. Mitchell, 1987, A doubled CO2
climate sensitivity experiment with a global climate model
including a simple ocean, J. Geophysical Res., 92(D11):13,315-13,343.
Wu, C., 1990, Land Use Map of China (1:1,000,000),
Science Press, Beijing. (in Chinese)
Wu, C., and H. Guo, 1994, Land Use of China,
Science Press, Beijing, p. 90. (in Chinese)
Xiao, X., D.S. Ojima, W.J. Parton, Z. Chen and D. Chen,
1995, Sensitivity of Inner Mongolia grasslands to global climate
change, Journal of Biogeography, 22:643-648.
Xiao, X., D.W. Kicklighter, J.M. Melillo, A.D. McGuire,
P.H. Stone and A.P. Sokolov, 1996a, Linking a global terrestrial
biogeochemistry model and a 2-dimensional climate model:
Implications for the global carbon budget, Tellus (in
press).
Xiao, X., J.M. Melillo, D.W. Kicklighter, A.D. McGuire,
P.H. Stone and A.P. Sokolov, 1996b, The relative roles of changes
in CO2 and climate to the equilibrium responses of net primary
production and carbon storage of the terrestrial biosphere, Global
Change Biology (in review).
Xiao, Q., W. Chen, Y. Sheng and L. Guo, 1996, Estimating
the net primary productivity in China using meteorological
satellite data, Acta Botanica Sinica, 38(1):35-39.
(in Chinese with English abstract)
Xiong, Y., and Q. Li (eds.), 1987, Soils of China,
Science Press, Beijing, p. 333. (in Chinese)
Zhang, X., 1993, A vegetation-climate classification
system for global change studies in China, Quaternary
Sciences, 2:157-169. (in Chinese with English
abstract)
Zhang, Y., and L. Liu, 1993, The potential impact of
climate change on vegetation distribution in Northwest China. In:
"Climate Change and Its Impact," Y. Zhang, P. Zhang, H.
Zhang and Z. Lin (eds.), Meteorology Press, Beijing, p. 178-193.
(in Chinese with English abstract)
Zhang, Y., and J. Song, 1993, The potential impact of
climate change on the vegetation in the Northeast China, ibid,
p. 194-204.
Zhang, Y., and W. Wang, 1993, The potential change of
ground surface air temperature under the condition of global
warming induced by CO2 doubling, ibid, p. 248-255.
Zhao, M., 1993, Impact of climate change on physical
geographical zones of China, ibid, p. 168-177.
Zhou, G., and X. Zhang, 1996, Study on climate-vegetation
classification for global change in China, Acta Botanica Sinica,
38(1):8-17. (in Chinese with English abstract)