![]()
--> Home Page | Report Series |
X. Xiao, J.M. Melillo, D.W. Kicklighter, A.D. McGuire, P.H. Stone and A.P. Sokolov
June 1996
View Report as PDF file (260 kB)
Contents
Increases in anthropogenic emissions of greenhouse gases (e.g., CO2, CH4, N2O) have resulted in an increase of the atmospheric abundance of these gases and consequently an increase of radiative forcing of climate (Houghton et al., 1995). Atmospheric general circulation models (GCMs) have indicated that the increase of radiative forcing from a doubled CO2 will change temperature, precipitation and cloudiness of the globe. According to a number of GCMs, equilibrium climate change simulations for a doubled CO2 will result in increases of global surface mean annual temperature between 1.5 degrees C and 4.5 degrees C (Mitchell et al., 1990). The GCMs also project large changes in magnitudes and spatial distributions of precipitation and cloudiness for a doubled CO2.
Changes in atmospheric CO2 concentration and climate (temperature, precipitation, solar radiation) are likely to have impacts on the structure and function of the terrestrial biosphere (Gates, 1985; Melillo et al., 1990). There are few field experiments on the responses of whole terrestrial ecosystems to elevated CO2 (Mooney et al., 1991) and to climate change, e.g., soil warming (Van Cleve et al., 1990; Melillo et al., 1995). At present, there is no full-factorial, long-term field experiment to examine the combined effects of changes in individual variables (CO2 level, temperature, precipitation and solar radiation) and their interactions on terrestrial ecosystems. Our understanding about the sensitivity of net primary production and carbon stock of the terrestrial ecosystems to a change in CO2 and changes in temperature, precipitation and cloudiness (solar radiation) projected by the climate models is limited to the results of modeling studies.
In earlier sensitivity studies that have used ecosystem models, investigators applied +1 degrees C, +2 degrees C, +4 degrees C warming and/or ±10%, ±20% increase of precipitation uniformly over the globe or continents to estimate the response of NPP (Esser, 1987, 1990; McGuire et al., 1993; Zhang, 1993) and carbon storage to climate change (McGuire et al., 1996a, 1995; Melillo et al., 1995; Schimel et al., 1994; Potter et al., 1993; Townsend et al., 1992; Jenkinson et al., 1991; Buol et al., 1990). Although this single factor sensitivity analysis approach has been useful for preliminary analyses of the effects of climate change on ecosystems, interpretation of the results have been limited. In reality, changes in climate variables will not be uniform across regions, and GCMs project large latitudinal and longitudinal variations in temperature and precipitation changes for doubled CO2. By not considering the covariations in climate variables, this approach has the largest inconsistency of the physical climate system and has not taken the effects of other abiotic controls on NPP and carbon storage into account. The interaction between a change in one abiotic factor and changes in other abiotic factors may play an important role in the responses of terrestrial ecosystems to climate change.
To better account for the physical consistency of climate change and the interactions among the driving variables, a number of studies have used the projected changes in temperature, precipitation and cloudiness from GCMs and elevated CO2 to simulate the potential impacts of climate changes and elevated CO2 on primary production and carbon storage of natural and managed ecosystems at global and continental scales (Melillo et al., 1993; Parton et al., 1995; Rosenzweig and Parry, 1994; VEMAP Members, 1995; Xiao et al., 1996). Both the magnitude and spatial distribution of responses of net primary production (NPP) and carbon storage vary among climate change predictions as estimated by GCMs for an particular ecosystem model. The responses of NPP and carbon storage also vary among various ecosystem models for a given climate change prediction (VEMAP Members, 1995). The differences in responses of NPP and carbon storage may be attributable to the concepts and formulations of the ecosystem models and climate models used in the studies. One of the earlier studies also showed that the response of net primary production of the terrestrial biosphere is affected strongly by the interaction between a change in atmospheric CO2 concentration and a change in climate (Melillo et al., 1993). Although this approach provides estimates of the overall responses of NPP and carbon storage to changes in the four driving variables, the relative contribution of a change in an individual variable and its interactions with changes of the other three variables to the overall responses of NPP and carbon storage have not been quantified.
Uncertainties in future concentrations of atmospheric CO2 and projected changes in temperature, precipitation and cloudiness by GCMs are large and differ among the four variables. Different ecosystems may have different responses to changes in CO2, temperature, precipitation and cloudiness. Quantification of the effect of a change in an individual driving variable, including its interaction with changes of other driving variables, on NPP and carbon storage will further our understanding of responses of ecosystems to climate change and how well ecosystem models represent ecosystem processes. In addition, we will gain a better understanding of the importance of the uncertainties in changes of climate variables estimated by GCMs on ecosystems.
In this study, we conduct a partial factorial experiment of model simulations, using a global biogeochemistry model (Terrestrial Ecosystem Model - TEM 4.0; McGuire et al., 1995a, 1996b) to quantify the relative importance of projected changes in temperature, precipitation, cloudiness and atmospheric CO2 concentration to the equilibrium responses of NPP and carbon storage of terrestrial ecosystems. We used two levels of atmospheric CO2 concentration (315 ppmv and 522 ppmv) and changes in temperature, precipitation and cloudiness projected by a 3-dimensional GCM (Geophysical Fluid Dynamic Laboratory - GFDL; Manabe and Wetherald, 1987; Wetherald and Manabe, 1988) and a 2-dimensional statistical-dynamical climate model (Land-Ocean climate model at Massachusetts Institute of Technology (MIT L-O); Sokolov and Stone, 1995). Both the 2-D MIT L-O climate model and TEM are linked in an integrated impact assessment framework of climate change at Massachusetts Institute of Technology (Prinn et al., 1996; Xiao et al., 1996). A comparison of the responses of primary production and carbon storage to climate change predictions between the 3-D GFDL GCM and the 2-D MIT L-O climate model provide insights into the importance of spatial resolutions, including longitudinal variations of climate change predictions, on estimating ecosystem responses to climate change (Xiao et al., 1996). To explore how ecosystem responses vary over different spatial scales, we examined the responses of NPP, reactive soil organic carbon and vegetation carbon across the scales of the globe, latitudinal bands and biomes.
The TEM (Raich et al., 1991; McGuire et al., 1992, 1993, 1995a, 1996a, 1996b) is a process-based ecosystem model that simulates important carbon and nitrogen fluxes and pools for various terrestrial ecosystems (Fig. 1, below). It runs at a monthly time step. Driving variables include monthly mean climate (precipitation, temperature and cloudiness), soil texture (sand, clay and silt proportion), elevation, vegetation and water availability. The water balance model of Vorosmarty et al., (1988) is used to generate hydrological input (e.g., potential evapotranspiration, soil moisture) for TEM. In this study, we used version 4.0 of TEM (McGuire et al., 1995a, 1996b). Here, we briefly review how TEM describes the influence of CO2, temperature, precipitation and cloudiness on primary production and carbon stocks.
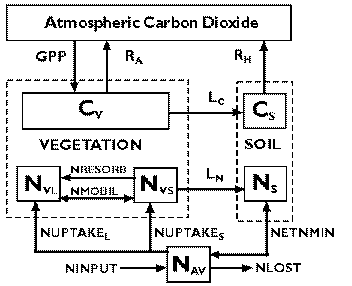
Figure 1. The Terrestrial
Ecosystem Model (TEM). The state variables are: carbon in
vegetation (CV); structural nitrogen in vegetation (NVS); labile
nitrogen in vegetation (NVL); organic carbon in soils and
detritus (CS); organic nitrogen in soils and detritus (NS); and
available soil inorganic nitrogen (NAV). Arrows show carbon and
nitrogen fluxes: GPP, gross primary productivity; RA, autotrophic
respiration; RH, heterotrophic respiration; LC, litterfall
carbon; LN, litterfall nitrogen; NUPTAKES, N uptake into the
structural N pool of the vegetation; NUPTAKEL, N uptake into the
labile N pool of the vegetation; NRESORB, N resorption from dying
tissue into the labile N pool of the vegetation; NMOBIL, N
mobilized between the structural and labile N pools of the
vegetation; NETNMIN, net N mineralization of soil organic N;
NINPUT, N inputs from the outside of the ecosystem; and NLOST, N
loss from the ecosystem.
Net primary production (NPP) is calculated as the difference between gross primary production (GPP) and plant respiration (RA). The flux GPP is calculated at each monthly time step as follows:
where Cmax is the maximum rate of C assimilation, PAR is photosynthetically active radiation, LEAF is leaf area relative to maximum annual leaf area, T is temperature, CO2 is atmospheric CO2 concentration, H2O is water availability, and NA is nitrogen availability.
The effect of CO2 and water availability on GPP are interrelated. The function f(CO2, H2O) is described by the hyperbolic relationship:
where Ci is the concentration of CO2 within leaves of the canopy and kc is the half-saturation constant for CO2 uptake by plants. The relationship between CO2 concentration inside stomatal cavities (Ci) and in the atmosphere (Ca) is directly proportional to relative moisture availability (Raich et al., 1991):
where Gv is a unitless multiplier that accounts for changes in leaf conductivity to CO2 resulting from changes in moisture availability, PET is potential evapotranspiration and EET is the estimated evapotranspiration. The flux PET is calculated as a function of mean air temperature and solar radiation (Jensen and Haise, 1963). The flux EET is equal to PET in wet months but is modeled as a function of rainfall, snowmelt recharge and a change of soil moisture in dry months (Vorosmarty, 1989).
The results from CO2-enrichment studies indicate that the response of plant productivity to doubled CO2 ranges from 20% to 50%, given adequate nutrients and water (Kimball, 1975; Gates, 1985; McGuire et al., 1995b). In TEM, the parameter kc (400 ppmv) has been chosen to increase f(CO2, H2O) by 37% for a doubling of atmospheric CO2 concentration from 340 ppmv to 680 ppmv, with canopy conductance equal to 1 (McGuire et al., 1992, 1993). It is important to note that the response of GPP to doubled CO2 is not a constant 37% for kc of 400 ppmv, because GPP calculation is also affected by f(NA), which represents the limiting effect of vegetation nitrogen status on GPP (McGuire et al., 1992, 1993; Melillo et al., 1993). Vegetation nitrogen status is determined by vegetation nitrogen uptake (NUPTAKE) and nitrogen from the labile nitrogen pool of vegetation (NMOBIL). The nitrogen down-regulation of GPP response to elevated CO2 in TEM is discussed elsewhere (McGuire et al., 1996b).
Increasing irradiance of PAR increases GPP hyperbolically in the following form: f(PAR) = PAR / (ki + PAR). A mean value of 314 J cm^-2 d^-1 for ki from published leaf studies is used and applied to entire leaf canopies independent of vegetation types (Raich et al., 1991). Cloudiness affects the amount of solar irradiance, including PAR, that reaches the canopy of vegetation.
The effect of air temperature on GPP is described by allowing f(T) to increase in a parabolic fashion to a grid-cell specific optimum temperature. Between the optimum temperature and a maximum vegetation-specific temperature constraint, f(T) is equal to 1.0. Above the maximum temperature constraint, f(T) declines rapidly to 0.0 (McGuire et al., 1996b). Air temperature also affects plant respiration (RA). The flux RA includes both maintenance respiration (RM) and construction respiration (RC). The flux RM increases logarithmically with temperature using a Q10 value that varies from 1.5 to 2.5 (McGuire et al., 1992). The flux RC is determined to be 20% of the difference between GPP and RM (Raich et al., 1991). Thus, changes in NPP are directly related to changes in CO2, temperature, precipitation and cloudiness.
In TEM, the amount of carbon stored in vegetation and soils is a balance between carbon fluxes into and out of these pools (Fig. 1). To attain equilibrium conditions in the simulation, annual fluxes out of a carbon pool must equal annual fluxes into the pool. Vegetation carbon increases with NPP, but decreases with litterfall. Although litterfall is simply modeled as a function of biomass with no independent climate and CO2 effects, annual litterfall must equal annual NPP to maintain equilibrium conditions in the simulation. Thus, climate and CO2 have an indirect effect on litterfall. Soil organic carbon increases with litterfall, but decreases with decomposition (i.e., heterotrophic respiration, RM). The flux RM represents decomposition of all organic matter in the ecosystem. It is modeled as a function of soil carbon (Cs), mean monthly air temperature (T), mean volumetric soil moisture and the gram-specific decomposition constant kd (Raich et al., 1991):
where MOIST is a parabolic function of volumetric soil moisture (McGuire et al., 1996b).
Thus, changes in heterotrophic respiration depend directly on changes in temperature and precipitation. Changes in CO2 and cloudiness also indirectly influence decomposition of soil organic matter by affecting litterfall and the resulting amount of soil organic matter to be decomposed.
In this study, we designed a partial factorial experiment (Table 1, below) to efficiently illustrate the combined effects of a change in an individual variable and its interactions with changes of other variables on the overall responses of NPP and carbon storage of the terrestrial biosphere to changes in CO2 and climate. We first ran TEM for contemporary climate with 315 ppmv CO2 as the baseline. Next, we ran TEM for complete climate change at 522 ppmv CO2 (see #1, #6 in Table 1), which is considered as the full response model for our comparison in this study. Third, we ran four TEM simulations (see #2-5, #7-10 in Table 1). In each of the four simulations, one of the four driving variable (CO2, temperature, precipitation, cloudiness) uses the baseline values (i.e., contemporary climate, 315 ppmv CO2), while the other three variables use values from the climate change predictions or elevated CO2. Eliminating a change in an individual variable in a simulation (e.g., D CO2) will result in losses of both the effect of the change in this variable (i.e., D CO2) and the effect of its interactions with changes of other variables, i.e., temperature (D CO2 x D T), precipitation (D CO2 x D P) and cloudiness (D CO2 x D C). This approach has the least inconsistency in physical climate system and includes the interactions among changes of three variables. Therefore, this subset of six model runs out of sixteen runs required in a full factorial experiment (4 factors at 2 levels) allows us to efficiently explore the relative contribution of a change in each of the four driving variables to the overall responses of primary production and carbon storage of the terrestrial biosphere. In the data analysis, we compare the results from the complete climate change at 522 ppmv CO2, to the four simulations that use the baseline values for one of the four driving variables. This comparison illustrates the combined effects of a change in an individual variable and its interactions with changes of the other three variables to the overall responses of primary production and carbon storage of terrestrial ecosystems.
To determine the relative contribution of the interaction between elevated CO2 and climate change to ecosystem responses, we compare the results among the following three simulations: (1) complete climate change at 522 ppmv CO2; (2) the scenario with no change in CO2 (complete climate change at 315 ppmv CO2); and (3) contemporary climate at 522 ppmv CO2, which is an additional TEM simulation to estimate the effect of elevated CO2 alone on primary production and carbon storage (Table 1). For examining the effects of changes in CO2 and climate at the global and biome scales, we may conceptualize that the responses of net primary production to complete climate change at 522 ppmv CO2 (D NPPD CO2, D climate) are represented by elevated CO2 (D CO2), climate change (D climate), and their interactions (D CO2 x D climate):
We can then calculate the effects of interactions on NPP as the following:
where D NPPD CO2 x D climate is the interaction between elevated CO2 and climate change, D NPPD CO2 is NPP response to contemporary climate at 522 ppmv CO2, and D NPPD climate is NPP response to complete climate change at 315 ppmv CO2. The same approach is used for vegetation carbon and soil carbon storage.
For global extrapolation, TEM uses spatial data sets that are gridded at a resolution of 0.5 degrees latitude by 0.5 degrees longitude. The global data sets include long-term average contemporary climate data from the Cramer and Leemans climate database (Cramer, personal communication), potential vegetation (Melillo et al., 1993), soil texture (FAO-UNESCO, 1971) and elevation (NCAR/Navy, 1984). The Cramer and Leemans climate data sets are an update of the Leemans and Cramer climate data sets (Leemans and Cramer, 1990). These spatial data sets contain 62,483 land grid cells, including 3,059 ice grid cells and 1,525 wetland grid cells. Geographically, the global data sets cover land areas from 56 degrees S to 83 degrees N. For climate change scenarios, we use climate outputs for 1xCO2 and 2xCO2 simulations from the 3-dimensional GFDL GCM (Weatherald and Manabe, 1988) and the 2-dimensional MIT L-O climate model (Sokolov and Stone, 1995). The 3-D GFDL GCM has a spatial resolution of 7.5 degrees (longitude) x 4.44 degrees (latitude). The 2-D MIT L-O climate model simulates the zonally averaged climate separately over land and ocean as a function of latitude and height (Yao and Stone, 1987; Stone and Yao, 1987, 1990; Sokolov and Stone, 1995). The 2-D model has 23 latitude bands, corresponding to a resolution of 7.826 degrees, and nine vertical layers. Detailed descriptions of the 2-D MIT L-O climate model for climate change predictions are documented in Sokolov and Stone (1995). The climate output of GFDL has been interpolated to 0.5 degrees x 0.5 degrees grid cells by applying a spherical interpolation routine to the data (Willmott et al., 1985). For the climate output of the 2-D MIT L-O climate model, we apply the zonally averaged data over land to all 0.5 degrees x 0.5 degrees grid cells within the latitudinal band.
Globally-averaged changes in climate variables
projected by the 3-D GFDL are in the middle of the range of
changes projected by a number of GCMs for doubled CO2. The
globally-averaged changes in climate variables estimated by the
2-D MIT L-O climate model for doubled CO2 are similar to GFDL,
although they operate at very different spatial domains.
Globally, the GFDL GCM projects a change of +4.0 degrees C for
mean annual temperature, +8.3% for annual precipitation and -0.7%
for mean annual cloudiness. The 2-D MIT L-O climate model
projects a change of +4.2 degrees C for mean annual temperature,
+11.5% for annual precipitation and -2.6% for mean annual
cloudiness. Latitudinal distributions in the changes of
temperature, precipitation and cloudiness between the GFDL and
MIT L-O have similar patterns (Fig. 2). In an earlier
study (Xiao et al., 1996), we compared the responses of NPP and
carbon storage to climate changes represented by two
3-dimensional GCMs (i.e., GFDL, Wetherald and Manabe, 1988;
Goddard Institute for Space Studies - GISS, Hansen et al., 1983,
1984) and the 2-D MIT L-O climate model (see Sokolov and Stone,
1995). The simulation results have shown that the global
responses of NPP and total carbon storage are generally similar
among the climate change predictions from the 3-D GCMs and the
2-D MIT L-O climate model.
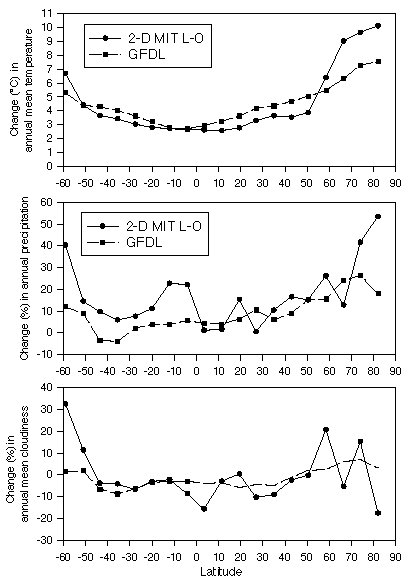
Figure 2. The zonal mean changes over land of
annual mean temperature, annual precipitation and annual mean
cloudiness between 1xCO2 and 2xCO2 simulations by the GFDL GCM
and the 2-D MIT L-O climate model along the latitudinal bands as
defined by the 2-D MIT L-O climate model. The outputs from the
GFDL are averaged over the same latitudinal bands as those of the
2-D MIT L-O climate model.
We used the following procedure to generate "future climate." First, we calculated absolute differences in monthly mean temperature, ratios in monthly precipitation and ratios in monthly mean cloudiness between the 2xCO2 simulation and the 1xCO2 simulation of each model. Then, we added the absolute differences in monthly mean temperature between the 2xCO2 and 1xCO2 simulations to the contemporary monthly temperature data; and multiplied the ratios in monthly precipitation and monthly mean cloudiness between the 2xCO2 and 1xCO2 simulations to the contemporary monthly precipitation and cloudiness data, respectively. For elevated CO2 level, we used an atmospheric CO2 concentration of 522 ppmv as corresponding to an effective CO2 doubling associated with the radiative forcing (Xiao et al., 1996). In this study, we ran TEM to equilibrium for each simulation. Therefore, the estimates of carbon and nitrogen fluxes and pool sizes apply only to mature, undisturbed vegetation and ecosystems. Effects of land use and management on carbon and nitrogen dynamics are not considered.
The TEM estimates global annual NPP to be 47.9 PgC yr^-1 for contemporary climate with 315 ppmv CO2. For complete climate change at 522 ppmv CO2, the global NPP response is +18.5% for the GFDL climate (see Tg, Pg, Cg, 522 in Table 2) and +17.8% for the 2-D L-O climate (see Tm, Pm, Cm, 522 in Table 3), respectively. Among the four simulations with the GFDL climate that use the baseline values for one variable, global NPP increases by 0.7% for the scenario with no change in CO2 (see Tg, Pg, Cg, 315 in Table 2); 7.7% for the scenario with no change in temperature (see Tcl, Pg, Cg, 522 in Table 2); 17.8% for the scenario with no change in precipitation (see Tg, Pcl, Cg, 522 in Table 2); and 19.0% for the scenario with no change in cloudiness (see Tg, Pg, Ccl, 522 in Table 2).
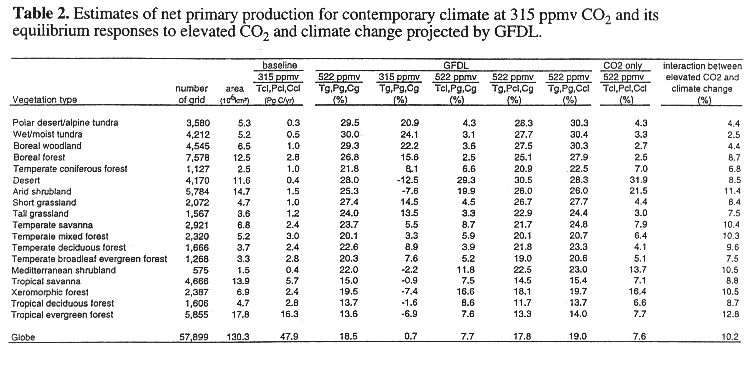
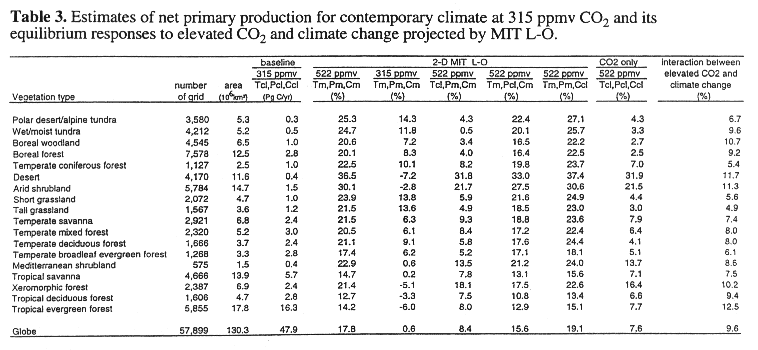
The difference in global NPP responses between the scenario of the complete climate change at 522 ppmv CO2 and one of the four scenarios that uses the baseline values for one variable indicates the combined effect of a change in an individual variable and its interactions with changes in other variables on the overall response of global NPP, i.e., the relative role of a change in an individual variable The difference ranges from -0.5% (18.5% - 19.0%) for the scenario with no change in cloudiness to 17.8% (18.5% - 0.7%) for the scenario with no change in CO2. Thus, the larger difference indicates a larger contribution of the change in CO2 to the overall NPP response. The results show that with the TEM, elevated CO2 contributes substantially, an increase in temperature contributes moderately, and changes in precipitation and cloudiness contribute little to the overall response of global NPP. Global NPP responses for the simulations using the 2-D L-O climate (Table 3) have patterns similar to those for the GFDL climate (Table 2).
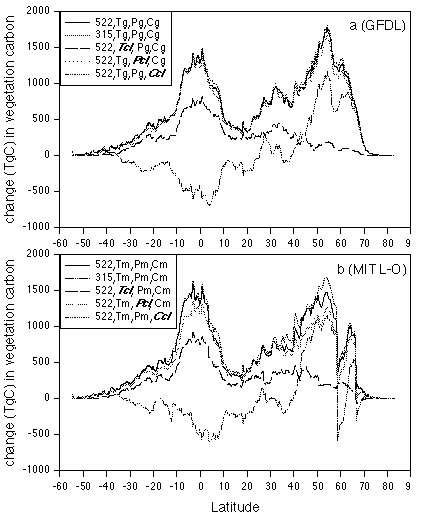
Along the 0.5 degree resolution latitudinal bands, NPP has a
bimodal distribution for complete climate change at 522 ppmv CO2
for both the GFDL climate (Fig. 3a) and the MIT L-O climate (Fig.
3b). There are large increases in both tropical regions and the
mid-latitude in the northern hemisphere. In the latitudinal bands
dominated by arid biomes, including the Sahara desert (i.e., 10
degrees N to 25 degrees N), the response of NPP to complete
climate change at 522 ppmv CO2 is more limited. For the scenario
with no change in CO2, NPP decreases in the tropical regions but
increases in the mid-latitude temperate zones (Fig. 3). For the
scenario with no temperature increase, NPP response is large in
tropical regions, but small in mid- to high- latitudes (Fig. 3).
The latitudinal distributions of NPP response for the scenario
with no change in precipitation and the scenario with no change
in cloudiness are similar to the latitudinal distribution of NPP
response to complete climate change at 522 ppmv CO2. Thus, the
changes in precipitation and cloudiness projected by the climate
models contribute little to the overall response of NPP along the
latitudinal bands.
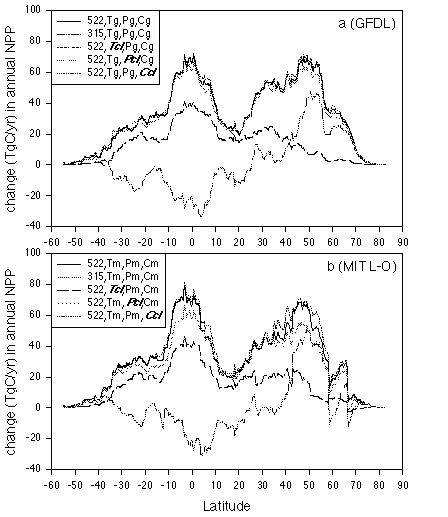
Figure 3. Latitudinal distributions of the responses
of net primary production to change in atmospheric CO2
concentration and changes in temperature, precipitation and
cloudiness projected by the 3-D GFDL and 2-D MIT L-O climate
models. The latitudinal bands have a 0.5 degree resolution. (a)
GFDL climate; (b) MIT L-O climate.
In TEM, NPP of mid- and high- latitude ecosystems is primarily limited by nitrogen availability. The results indicate that the response of NPP in these regions is controlled primarily by temperature increase and secondarily by elevated CO2. Increases of temperature and precipitation in mid- and high- latitudes enhance decomposition of soil organic matter. As a result, more nitrogen is released from soils to be available for plant uptake, and thus plant production increases. The changes in temperature estimated by the climate models have a larger effect on the decomposition of soil organic matter than the predicted changes in precipitation. In the tropic regions, NPP of moist tropical ecosystems is generally not limited by nitrogen availability. The response of NPP in tropical regions is controlled primarily by elevated CO2 and secondarily by temperature increase. Although NPP in arid regions (i.e., 10 degrees N to 25 degrees N) is limited by water availability, the NPP responses for the scenario with no change in precipitation indicate that the effect of projected changes in precipitation is small in arid regions. Temperature also has a small effect on NPP response in arid regions. Elevated CO2 appears to account for most of the NPP response in the arid regions as a result of enhanced water use efficiency. The NPP response for the scenario with no change in cloudiness is slightly larger than the NPP response to complete climate change at 522 ppmv CO2 (Fig. 3). The degree of cloudiness in the contemporary climate is greater than that projected for the future by the GFDL GCM and the MIT L-O climate model. In TEM, lower cloudiness increases the amount of solar radiance, including PAR. Enhanced PAR has the potential to increase GPP and NPP. Enhanced solar radiation also increases PET. High PET may reduce soil moisture and thus increase water stress of plant to potentially decrease GPP and NPP. The results indicate that the effect of enhanced radiation-induced water stress is larger than the effect of increased PAR.
The latitudinal distributions of NPP responses to the GFDL climate are similar to the latitudinal distributions of NPP responses to the MIT L-O climate, except at high latitudes. The largest differences in NPP responses occur within the 50.5 - 58.5 degrees N and 66.5 - 74.0 degrees N latitudinal bands, where the MIT L-O climate model projects relatively larger increases in temperature and cloudiness than the GFDL GCM (Fig. 2). The comparison of NPP responses among the scenario with no temperature change, the scenario with no cloudiness change and the scenario with precipitation change indicates that the differences in NPP responses at high latitudes between the GFDL climate and MIT L-O climate are primarily due to larger increases in temperature projected by the MIT L-O climate model (Fig. 2).
At the biome scale, the response of NPP varies among the 18 vegetation types. NPP increases substantially in all biomes for complete climate change at 522 ppmv CO2, ranging from +13.6% in tropical evergreen forest to 30.0% in wet/moist tundra for the GFDL climate (Table 2) and from 12.7% in tropical deciduous forest to 36.5% in desert for the MIT L-O climate (Table 3). For the scenario with no change in CO2, climate change projected by the GFDL results in a decrease of NPP in tropical ecosystems and arid ecosystems ranging from -12.5% in desert to -0.9% in tropical savanna, but an increase of NPP for the other biomes, ranging from +3.3% in temperate mixed forest to +24.1% in wet/moist tundra (Table 2). The results indicate that percent response of NPP to a change in CO2 is most sensitive in deserts and the least sensitive in wet/moist tundra. Elevated CO2 significantly increases water use efficiency of plants in arid lands. Although percent NPP responses in moist tropical ecosystems are moderate, the absolute changes of NPP in tropical ecosystems are substantial, because NPP per unit area in tropical ecosystems is much larger than it is in deserts and arid shrublands.
For the scenario with no change in temperature, NPP increases for the 18 biomes, ranging from 2.5% in boreal forest to 29.3% in desert for the GFDL-q climate (Table 2), and from 0.5% in wet/moist tundra to 31.8% in desert for the MIT L-O climate (Table 3). The NPP response for the scenario with no temperature increase is much smaller than the NPP response for complete climate change with 522 ppmv CO2, except for arid ecosystems (e.g., desert, arid shrubland) where water is the dominant limiting factor and soil organic matter is low (Table 2). Thus, NPP of most ecosystems is sensitive to changes in temperature. The NPP response to changes in temperature estimated by GFDL is the most sensitive in boreal forest and the least sensitive in desert. The NPP responses for the scenario with no change in precipitation and the scenario with no change in cloudiness are similar to the NPP responses to complete climate change at 522 ppmv CO2 for all 18 biomes. These results indicate that projected changes in precipitation and cloudiness by the climate models have little effect on NPP at the biome scale. The responses of NPP for the scenario with no change in cloudiness are slightly higher than NPP responses to complete climate change at 522 ppmv CO2. As contemporary climate has slightly higher cloudiness, the results indicate that the effect of enhanced radiation-induced water stress to plants is larger than the effect of increasing PAR for all biomes.
The interaction between elevated CO2 and climate change contributes about 10.2% to the overall response (18.5%) of global NPP to complete climate change at 522 ppmv CO2 in the GFDL climate (Table 2). Similarly, the interaction between elevated CO2 and climate change contributes 9.5% to the overall response (17.8%) of global NPP to complete climate changes at 522 ppmv CO2 in the MIT L-O climate (Table 3). Globally, the interaction between elevated CO2 and climate change has a larger contribution to the overall response of NPP than the effect of elevated CO2 alone. In conjunction with the analyses on the relative role of changes in CO2, temperature, precipitation and cloudiness in the previous section, the results show that the interaction between elevated CO2 and climate change is primarily determined by the interaction between elevated CO2 and temperature change. Higher temperature results in enhanced decomposition of soil organic matter (see Eq. 4) to make more nitrogen available in soils for plant uptake. Both higher temperature and elevated CO2 may increases GPP (see Eqs. 1, 2, 3). Therefore, NPP increases substantially for the scenarios including both elevated CO2 and temperature change, because more carbon (CO2) and nitrogen are available.
At the biome scale, the interaction between elevated CO2 and climate change is important but its contribution to the overall NPP response varies among the biomes (Tables 2, 3). In tropical evergreen forest, the interaction between elevated CO2 and climate change plays a dominant role and is much larger than both the effect of climate change and the effect of elevated CO2 alone. In boreal forests, climate change plays the largest role, the interaction between elevated CO2 and climate change an intermediate role, and elevated CO2 the least role. In contrast, the interaction between elevated CO2 and climate change in dry ecosystems (desert, arid shrubland, xeromorphic forest) is much smaller than the effect of elevated CO2 alone. The contribution of the interaction between elevated CO2 and climate change to the overall NPP response of the 18 biomes varies between the 3-D GFDL climate and 2-D MIT L-O climate, especially for high latitude ecosystems. Geographical distributions of climate changes differ between the climate models and affect the contributions of the interaction between elevated CO2 and climate change to the overall NPP responses at larger spatial scale.
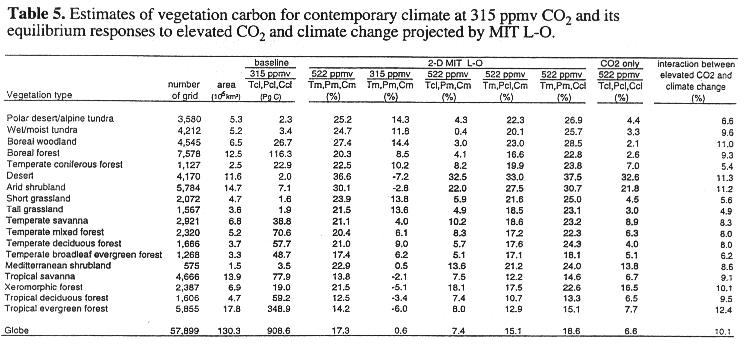
The TEM estimates global vegetation to be 909 PgC for contemporary climate with 315 ppmv CO2. Global vegetation carbon increases substantially for the complete climate change with 522 ppmv CO2: +18.3% (166 PgC) for the GFDL climate and +17.3% (157 PgC) for the MIT L-O climate.
The responses of vegetation carbon (Tables 4, 5, Fig. 4) have patterns similar to the NPP responses at the scales of latitudinal bands and biomes. An increase in NPP results in an increase in vegetation carbon. The responses of vegetation carbon are similar between the GFDL climate and MIT L-O climate, except at high latitudes (Fig. 4). The largest differences in vegetation carbon response between the GFDL climate and the MIT L-O climate occur within the 50.5 - 58.5 degrees N and 66.5 - 74.0 degrees N latitudinal bands, where the MIT L-O climate model projects relatively larger increases in temperature and cloudiness than the GFDL GCM (Fig. 2).
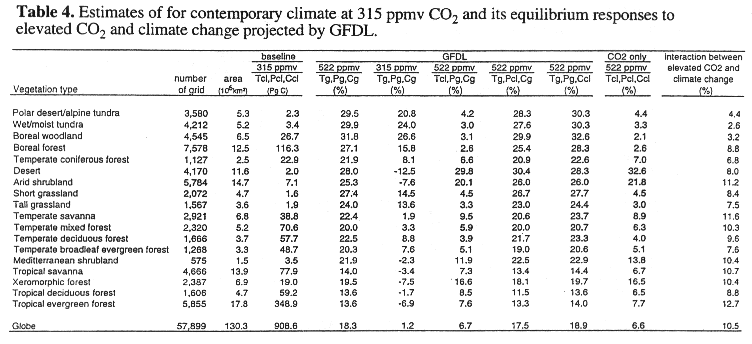
Version 4.0 of TEM defines a reactive soil carbon pool that excludes biologically "inert" soil organic matter. The TEM estimates that global reactive soil organic carbon at equilibrium is 750 PgC for contemporary climate with 315 ppmv CO2, which is about 50% of the 1500 PgC estimated by several inventories of soil organic carbon up to 1-meter depth (Schlesinger, 1977; Post et al., 1982; Eswaran et al., 1993).
For complete climate change at 522 ppmv CO2, global reactive soil organic carbon decreases moderately by -4.0% (30 PgC) for the GFDL climate (Table 6) and by -5.6% (42 PgC) for the MIT L-O climate (Table 7). For the scenario with no change in CO2, global reactive soil organic carbon decreases substantially, i.e., -15.0% (112 PgC) for the GFDL climate and -17.2% (129 PgC) for the MIT L-O climate. For the scenario with no change in temperature, global reactive soil organic carbon increases moderately by +4.8% (36 PgC) for the GFDL climate and +5.0% (38 PgC) for the MIT L-O climate, respectively. Global reactive soil organic carbon decreases moderately for the scenario with no change in precipitation and the scenario with no change in cloudiness (Tables 6, 7).
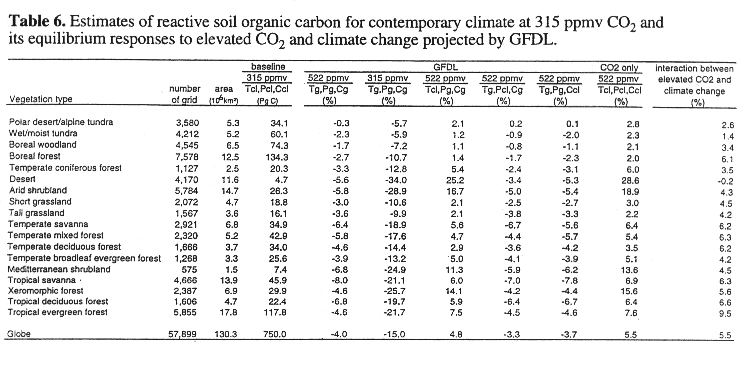
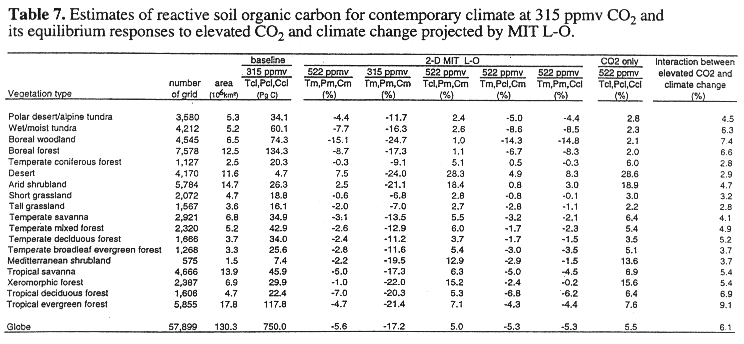
A comparison of the responses of soil organic carbon between the scenario of complete climate change at 522 ppmv CO2 and the four scenarios that use baseline values for one of the four driving variables indicates that temperature increase and elevated CO2 contribute the most to the response of reactive soil organic carbon. The changes in precipitation and cloudiness contribute little to the loss of reactive soil organic carbon. In TEM, an increase in temperature enhances decomposition of soil organic matter, resulting in an decrease of soil organic carbon. Elevated CO2 increases NPP, which then increases litterfall. More input of litterfall to soils would increase soil organic carbon. Globally, the responses of reactive soil organic carbon in the MIT L-O climate are slightly larger than those in the GFDL climate. This difference in response is attributable to relatively larger temperature increases projected by the MIT L-O climate model.
Along the 0.5 degree resolution latitudinal
bands, changes in reactive soil organic carbon vary slightly for
the complete climate change at 522 ppmv CO2, with relatively
larger decreases in mid- latitudes. Reactive soil organic carbon
has the largest decrease for the scenario with no change in CO2.
In contrast, reactive soil organic carbon increases in all
latitudinal bands for the scenario with no change in temperature.
The responses of reactive soil organic carbon for the scenario
with no change in precipitation and the scenario with no change
in cloudiness are similar to that for the complete climate change
at 522 ppmv CO2 (Fig. 5). In general, the loss of reactive soil
organic carbon is relatively larger in mid- to high- latitudes in
the northern hemisphere than in tropical regions for the
scenarios with temperature increase (Fig. 5). For doubled CO2,
both the GFDL and MIT L-O climate models project that increases
in temperatures are larger in the mid- and high- latitudes than
in tropical regions (Fig. 2). Latitudinal distributions of
reactive soil organic carbon for the GFDL climate are similar to
those for the MIT L-O climate, except at high latitudes of the
northern hemisphere, where loss of soil organic carbon is
substantially larger for the MIT L-O climate than for the GFDL
climate (Fig. 5). This is attributable to the relatively larger
temperature increases at high latitudes projected by the MIT L-O
climate model (Fig. 2). The projected temperature increases at
high latitudes by the MIT L-O climate model are similar to those
for the UKMO climate (Wilson and Mitchell, 1987).
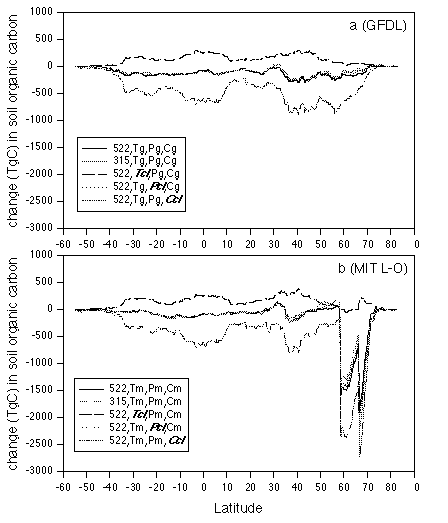
Figure 5. Latitudinal distributions of the
responses of reactive soil organic carbon to change in
atmospheric CO2 concentration and changes in temperature,
precipitation and cloudiness projected by the 3-D GFDL and 2-D
MIT L-O climate models. The latitudinal bands have a 0.5 degree
resolution. (a) GFDL climate; (b) MIT L-O climate.
At the biome scale, reactive soil organic carbon decreases in the 18 biomes for complete climate change at 522 ppmv CO2, ranging from -0.3% in polar desert/alpine tundra to -8.0% in tropical savanna for the GFDL climate (Table 6), and from -0.3% in temperate coniferous forest to -15.1% in boreal woodlands for the MIT L-O climate (Table 7). All biomes lose soil organic carbon for the scenario with no change in CO2, but accumulate soil organic carbon for the scenario with no change in temperature (Table 6). The dry ecosystems (desert, arid shrubland, xeromorphic forest and Mediterranean shrubland) gain more soil organic carbon than the other ecosystems in the scenario with no change in temperature, because of reduced water stress on production in the dry regions. A comparison of responses of soil organic carbon between the scenario of complete climate change at 522 ppmv CO2 and the scenario with no precipitation change indicates that the increase of precipitation projected by the climate models slightly enhances decomposition of reactive soil organic carbon for the 18 biomes. A decrease of cloudiness has a minor effect on reactive soil organic carbon. Large differences in loss of reactive soil organic carbon between the GFDL climate and the MIT L-O climate occur in higher latitude ecosystems, for instance, boreal forest, boreal woodland, wet/moist tundra (Tables 6, 7). Again, this is attributable to a larger temperature increase in high latitudes projected by the 2-D MIT L-O climate model (Fig. 2).
At the global scale, the response of reactive soil organic carbon to complete climate change at 522 ppmv CO2 is dominated by the loss of soil organic carbon caused by the temperature increase (Tables 6, 7). The compensating effects of elevated CO2 and the interaction between elevated CO2 and climate change reduce the amount of soil organic carbon that could potentially be lost from terrestrial ecosystems. Globally, the contribution of the interaction between elevated CO2 and climate change to the overall response of reactive soil organic carbon under complete climate change at 522 ppmv CO2 is 5.5% for the GFDL climate and 6.1% MIT L-O climate, respectively, which are equal or slightly larger than the contribution (5.5%) from the main effect of elevated CO2 (Tables 6, 7). The interaction between elevated CO2 and climate change represents the effect of enhanced NPP and litterfall on soil organic carbon pool caused by elevated CO2 and temperature increase.
At the biome scale, the interaction between elevated CO2 and climate change has different roles in the overall response of soil organic carbon among the 18 biomes (Tables 6, 7). In dry biomes (desert, arid shrubland, xeromorphic forest, Mediterranean shrubland), the interaction between elevated CO2 and climate change plays a much smaller role than the main effect of elevated CO2. In contrast, the interaction between elevated CO2 and climate change in tropical evergreen forest and boreal forest play a larger role than the main effect of elevated CO2. This is related to the fact that NPP in the dry biomes is primarily controlled by water availability and the amounts of soil organic matter in the dry biomes are much smaller than tropical evergreen forest and boreal forest (Tables 6, 7). The contribution of the interaction between elevated CO2 and climate change varies between the 3-D GFDL and the 2-D MIT L-O climate model. Therefore, the differences in the geographical distributions of climate change among climate models does affect the relative contribution of the interaction between elevated CO2 and climate change to the overall responses of reactive soil organic carbon at larger spatial scales.
The TEM results show that elevated CO2 and temperature change contribute large proportions to the overall responses of NPP and carbon storage to changes in CO2 and climate, while precipitation and cloudiness changes contribute small proportions. This is attributable to the magnitude of changes in CO2, temperature, precipitation and cloudiness projected by the GCMs. In an earlier study that used alternative input data sets of contemporary climate, solar radiation and soil texture for the conterminous United States to drive TEM (Pan et al., 1996), results show that the differences in NPP estimates depend in part on the magnitude of differences among the input data sets. The projected changes in global annual mean temperature, annual precipitation and annual mean cloudiness are similar between the GFDL and MIT L-O climate models. Other GCMs have also projected similar magnitudes of climate change for doubled CO2. For example, the GISS GCM projected +4.2 degrees C in global mean annual temperature, +11.0% in annual precipitation and -3.4% in mean annual cloudiness (Hansen et al., 1983, 1984). The OSU GCM projected +2.8 degrees C in global mean annual temperature, +7.8% in annual precipitation and -3.4% in mean annual cloudiness (Schlesinger and Zhao, 1989). Major differences among GCMs occur in spatial patterns of projected changes of temperature, precipitation and cloudiness, especially in precipitation (Cramer and Leemans, 1993). As shown in this study, the responses of NPP, vegetation carbon and reactive soil organic carbon are slightly different between the 3-D GFDL climate and the 2-D MIT L-O climate across the scales of the globe, latitudinal bands and biomes, except at high latitudes.
This study indicates that CO2 fertilization plays an important role in primary production and the global carbon budget. This is consistent with previous analyses using the TEM (Melillo et al., 1993, 1995). A number of studies have suggested that CO2 fertilization is one of the major mechanisms that account for the global carbon budget (Gifford, 1993). For the global carbon budget in the decade of 1980-1989, approximately 1.8 PgC/yr of anthropogenic emissions of carbon cannot be balanced with known carbon sinks and sources (Siegenthaler and Sarmiento, 1993). The estimates of the effect of global CO2 fertilization for the 1980s are ~1.2 PgC/yr (Rotmans and den Elzen, 1993), <1.5 PgC/yr (Friedlingstein et al., 1995), and 1 PgC/yr or less (Schimel, 1995). The estimate of the effect of CO2 fertilization on carbon storage in 1990 is about 1 PgC/yr (Melillo et al., 1996). However, these studies have not considered the possible effects of interactions among changes in CO2 and climate variables on carbon storage. Our results show that CO2 fertilization is influenced by climate change across the scales of the globe, latitudinal bands and biomes. At the global scale, the interaction between elevated CO2 and climate change has approximately an equal effect on the responses of NPP and carbon storage as the effect of elevated CO2 alone.
Our study also indicates that temperature change estimated by the climate models play an important role in responses of NPP and carbon stocks. The changes in annual mean temperature estimated by the GFDL and MIT L-O climate models for doubled CO2 are large, e.g., globally +4.0 degrees C by GFDL and +4.2 degrees C by MIT L-O. Increased temperature affects plant photosynthesis, plant respiration and water availability in soils. Increased temperature also affects the rates of temperature-dependent soil processes, e.g., decomposition and net nitrogen mineralization. In a soil warming experiment at the Harvard Forest site in Massachusetts (Melillo et al., 1995), soil warming of +5 degrees C resulted in a significant increase of carbon flux from the soils to atmosphere at the order of 56% (4000 kgC ha^-1 yr^-1) for the first year of the experiment and 13% (900 kgC ha^-1yr^-1) for the second year of the experiment. Soil warming also results in a doubling of net nitrogen mineralization rate in the forest floor and mineral soils for the first and second years of the experiment (Melillo et al., 1995). The responses of NPP and carbon storage in our study are closely related to enhanced decomposition of soil organic matter, nitrogen availability in soils and plant nitrogen uptake, as the result of climate change.
Although the GFDL and MIT L-O climate models project a moderate increase of annual precipitation for doubled CO2 globally (i.e., +8.3% for GFDL and +11.5% for MIT L-O), the changes in precipitation made small contributions to the responses of NPP and carbon storage. The difference in global NPP responses between the scenario of complete climate change at 522 ppmv CO2 and the scenario with no change in precipitation indicated that global NPP decreases 0.7% (0.34 PgC/yr) for the GFDL climate and 2.2% (1.10 PgC/yr) for the MIT L-O climate, because of lower precipitation (see Tables 2, 3). The results of this study showed that changes in temperature made a much larger contribution to the responses of NPP and carbon storage than changes in precipitation. In a simulation study that examines the response of NPP to climate change for 31 grassland sites worldwide, using the Century biogeochemistry model (Parton et al., 1995), changes in total plant production are shown to be more correlated to changes in temperature than to changes in precipitation.
Both the GFDL and MIT L-O climate models estimated a small change in cloudiness for doubled CO2, e.g. a decrease in globally averaged annual mean cloudiness (-0.7% for GFDL and -2.6% for MIT L-O). Lower cloudiness results in higher total solar radiation and photosynthetically active radiation. Higher solar radiation may enhance water stress of plants through an increase in potential evapotranspiration, which may results in lower net primary production. The difference in global NPP responses between the scenario of complete climate change at 522 ppmv CO2 and the scenario with no change in cloudiness indicated that global NPP increases 0.5% (0.24 PgC/yr) for the GFDL climate and 1.3% (0.62 PgC/yr) for the MIT L-O climate, because of higher cloudiness (see Tables 2, 3). Thus, a small change in cloudiness results in a small response of NPP and carbon storage of the terrestrial biosphere. The results are consistent with an earlier study that examined the effects of three alternative solar radiation data sets from the contemporary climate for the conterminous U.S. (Pan et al., 1996). In that study, solar radiation derived from the Cramer and Leemans climate database and the VEMAP project are 32% and 60% higher than the solar radiation derived from Hahn's cloudiness data, respectively. The estimates of annual NPP for the conterminous U.S. using solar radiation data from the Cramer and Leemans database and from the VEMAP project are about 8% and 10% lower than the NPP estimate using solar radiation data derived from the Hahn's cloudiness data (Pan et al., 1996).
In addition to cloudiness, aerosols also influence the amount of solar radiation that reaches the earth's surface. Aerosols from the eruption of Mt. Pinatubo in June 14-15, 1991 reduced solar irradiance by 4% and decreased temperature by about 0.5 degrees C within the period of summer 1991 to summer 1992 (Blumthaler and Ambach, 1994). After the volcanic eruption of Mt. Pinatubo, the atmospheric CO2 concentration began to drop in mid-1991 and decreased by 1.5 ppmv at Mauna Loa in May 1993 (Sarmiento, 1993). A large net sink of 2.5 PgC for this period, or 1.5 PgC/yr, is needed to account for the change in atmospheric CO2 concentration, and the 13C data seems to suggest that this carbon sink is mostly terrestrial (Sarmiento, 1993). Lower-than-normal temperatures may have caused net accumulation of carbon in a mature tropical rain forest in 1992-1993 (Grace et al., 1995). Given the sensitivity of carbon storage to a change in cloudiness, the reduction in solar radiation caused by the eruption may also result in a significant net terrestrial sink of carbon. However, the effects of aerosols was not included in the GFDL and 2-D MIT L-O climate models, and clouds are poorly represented in GCMs (Mitchell et al., 1989). Improving in projection of changes in cloudiness and aerosols by combined atmospheric chemistry models and GCMs will certainly be helpful in reducing uncertainty in responses of NPP and carbon storage to climate change.
In an earlier study on net primary production and carbon storage in the conterminous U.S. (VEMAP Members, 1995), TEM simulations indicated that the contributions from the interaction between climate change and elevated CO2 to the continental NPP responses range from 8% in the OSU climate to 19% in the UKMO climate. Although their analysis indicates the importance of the interaction between a change in CO2 and a change in climate, the relative role of a change in an individual climate variable and its interactions with changes in the other three driving variables was not quantified. However, the results presented in this paper show that interactions among changes in CO2 level and climatic variables contribute significantly to the equilibrium responses of net primary production and carbon storage. Furthermore, the results suggest that the interaction between a change in CO2 and a change in temperature is the most significant, given the magnitudes of changes in CO2 and climate variables projected by the climate models.
In TEM, the interaction between elevated CO2 and increased temperatures that influences NPP and carbon storage is caused by enhanced plant N uptake. As mentioned earlier, higher temperatures enhance decomposition of soil organic matter, resulting in decreases of soil carbon stocks. The rate of net nitrogen mineralization in soils increases with enhanced decomposition of soil organic matter. Mineralized nitrogen released from soil organic matter is assimilated by plants. Soil organic matter has low C/N ratio (10 - 20), while plant tissues have high C/N ratio (40 - 200). Thus, the shift of nitrogen from soils to vegetation allows substantial increase in vegetation carbon. Higher temperature also increases the rate of plant uptake of carbon (CO2) and nitrogen (Raich et al., 1991). Therefore, for complete climate change at 522 ppmv CO2, ecosystems are supplied with more mineralized nitrogen and carbon (CO2) resources, which results in large increases in NPP and total carbon storage (vegetation carbon plus reactive soil organic carbon, i.e., +137 PgC for the GFDL climate and +115 PgC for the MIT L-O climate).
Limited information is available on how the interactions among changes in CO2 and climate variables affect the carbon and nitrogen cycles of the terrestrial ecosystem, as there are few factorial field experiments for whole ecosystems. In a field study for a tundra ecosystem, which used two levels of atmospheric CO2 level (ambient CO2, 680 ppmv CO2) and temperature (ambient temperature, +4 degrees C temperature above ambient), the combination of elevated CO2 and temperature increase resulted in an increase of net carbon storage that lasted for the three years of observation (Oechel and Riechers, 1986, 1987). The interactions among temperature, CO2 and nutrient availability are key controlling factors in the responses of the tundra site (Melillo et al., 1990).
The analyses in this study are based on equilibrium or steady-state simulations of the TEM for potential natural vegetation. The results show that elevated CO2 and temperature increases projected by the climate models play a major role in the equilibrium responses of primary production and carbon storage of the terrestrial biosphere. The responses of NPP and carbon storage to the projected changes in CO2 and climate vary at the scales of the latitudinal bands and biomes. In addition, the results show that the interactions among changes in CO2 and climate variables, especially the interaction between elevated CO2 and temperature increase, may be important in the response of NPP and carbon storage. The relative role of the interaction between elevated CO2 and climate change also varies by biome.
Our results also imply that the time courses of changes in atmospheric CO2 concentration and climate are critical to the response of the terrestrial biosphere. In the next century, there is large uncertainty in anthropogenic emissions of CO2 from fossil fuel combustion, which will result in different rates of change in atmospheric CO2 concentration. Consequently, there are large uncertainties in the rate of climate change over time. The trajectory of changes in CO2 and climate in the next century may have significant impacts on the structural and functional responses of the terrestrial biosphere. The large interannual variation in climate is one of several processes that account for the imbalance of the global carbon budget in the last few decades (Dai and Fung, 1993). In future studies, we will conduct transient simulations of TEM to track both the path and magnitude of the responses of NPP and carbon storage to changes in CO2 and climate over time from the last century to the next century. Transient simulation of TEM will also allow us to incorporate the feedback of the terrestrial biosphere to the atmosphere (e.g., net carbon flux between lands and atmosphere) in the studies of land-atmosphere interactions (Melillo, 1994) and integrated assessment of climate change (Prinn et al., 1996; Xiao et al., 1996).
Changes in land use also affect terrestrial carbon storage and the global carbon cycle (Houghton and Skole, 1990). As the results of this study are based on potential vegetation, we have not considered the direct effects of human activities on NPP and carbon storage of the terrestrial ecosystems. However, the results presented in this study provide a baseline for us to assess the effects of changes in land use and land cover on the terrestrial carbon budget in future studies.
As shown in the comparisons between the 3-D GFDL climate and 2-D MIT L-O climate, the relative role of the individual climate variables and the interactions among changes in CO2 and climate variables to the overall response of NPP and carbon storage at larger spatial scales is affected by the geographical distributions of the projected changes in climate variables. There still are large uncertainties in the magnitudes and spatial distributions of changes of temperature, precipitation and cloudiness, as estimated by GCMs. Several climate modeling groups have begun to incorporate the effect of aerosols from anthropogenic sources on radiative forcing of climate for projections of future climate. Incorporation of the effect of aerosols into climate models would reduce the magnitude of temperature increases projected by climate models. Further improvement in climate models will reduce uncertainties in the magnitude and spatial distribution of climate change, and consequently uncertainties in responses of the terrestrial biosphere.
In a recent model comparison study (VEMAP Members, 1995), the estimates of the responses of NPP and carbon storage in the conterminous U.S. to climate change and elevated CO2 differ among the three biogeochemistry models. Although TEM has indicated that the interaction between elevated CO2 and climate change has an important effect on NPP and carbon storage, the results of the other models have indicated that this interaction is not important (VEMAP Members, 1995). Unfortunately, there is not enough information from field studies to confirm or deny the hypothesis that the interaction between elevated CO2 and climate change has an important effect on NPP and carbon storage of the terrestrial ecosystems. Therefore, long-term partial or full factorial field experiments at the whole ecosystem level are critically needed to quantify interactions among key driving variables of the terrestrial ecosystems, which will provide important information for development and validation of ecosystem models.
The TEM model and the 2-D MIT L-O climate model are component models of the modeling framework for the integrated assessment of global change in the Joint Program on Science and Policy of Global Change at the Massachusetts Institute of Technology (MIT). This study was supported by MIT (CE-S-462041), DOE NIGEC (No:901214-HAE) and NASA EOS (NAGW-2669). We thank Ronald G. Prinn, Henry D. Jacoby, Richard S. Eckaus, Yude Pan and John Helfrich for valuable discussions and comments.
Bazzaz, F.A. (1990) The response of natural
ecosystems to the rising global CO2 levels. Annual Review of
Ecology and Systematics, 21:167-196.
Blumthaler, M. and W. Ambach (1994) Changes in solar
radiation fluxes after the Pinatubo eruption. Tellus, 46B:76-78.
Buol, S.W., Sanchez, P.A., Kimble, J.M., and S.B. Weed
(1990) Predicted impact of climate warming on soil properties and
use. In: "Impact of Carbon Dioxide, Trace Gases, and Climate
Change on Global Agriculture," B.A. Kimball et al. (eds.),
ASA Spec. Publ., 53:71-82.
Cramer, W.P. and R. Leemans (1993)
Assessing impacts of climate change on vegetation using climate
classification systems. In: "Vegetation Dynamics and Global
Change," A.M. Solomon and H.H. Shugart (eds.), Chapman &
Hall, New York. p. 190-217.
Dai, A. and I.Y. Fung (1993) Can climate
variability contribute to the "missing" CO2 sink? Global
Biogeochemical Cycles, 7(3):599-609.
Esser, G. (1987) Sensitivity of global
carbon pools and fluxes to human and potential climate impact. Tellus,
39B:245-260.
Esser, G. (1990) Modeling global terrestrial sources and
sinks of CO2 with special reference to soil organic matter. In:
"Soils and Greenhouse Effect," A.F. Bouwman (ed.), John
Wiley, New York, p. 247-262.
Eswaran, H., Van Der Berg, E. and P. Reich (1993) Organic
carbon in soils of the world. Soil Science Society of America
Journal, 57:192-194.
FAO-UNESCO (1971) Soil map of the
world, 1:5,000,000. UNESCO, Paris, France. Digitization (0.5
degree resolution) by Complex Systems Research Center, University
of New Hampshire, Durham, and modifications by Marine Biological
Laboratory, Woods Hole.
Friedlingstein, P., Fung, I., Holland, E., John, J.,
Brasseur, G., Erickson, D. and D. Schimel (1995) On the
contribution of the biosphere CO2 fertilization to the missing
sink. Global Biogeochemical Cycles, 9(4):541-556.
Gates, D.M. (1985) Global biospheric
response to increasing atmospheric carbon dioxide concentration.
In: "Direct Effect of Increasing Carbon Dioxide on
Vegetation," B.R. Strain and J.D. Cure (eds.), DOE/ER-0238.
United States Department of Energy, Washington, D.C., p. 171-184.
Gifford, R.M. (1993) Implications of CO2 effects on
vegetation for the global carbon budget. In: "The Global
Carbon Cycle," M. Heimann (ed.), Proceedings of the NATO
Advanced Study Institute, II Ciocco, Italy, 8-20 September, 1991.
Springer-Verlag, New York, p. 159-199.
Hansen, J., Russell, G., Rind, D., Stone,
P., Lacis, A., Lebedeff, S., Ruedy, R. and L.Travis (1983)
Efficient three dimensional global models for climate studies:
Model I and II. Monthly Weather Review, 111:609-662.
Hansen, J., Lacis, A., Rind, D., Russell, G., Stone, P.,
Fung, I., Ruedy, R. and J. Lerner (1984) Climate sensitivity:
Analysis of feedback mechanisms. In: "Climate process and
Climate Sensitivity," J.E. Hansen and T. Takahashi (eds.),
Geophysical Monograph 29, Maurice Ewing series 5, American
Geophysical Union, Washington, D.C., p. 130-163.
Houghton, R.A. and D.L. Skole (1990) Changes in the global
carbon cycle between 1700 and 1985. In: "The Earth as
Transformed by Human Action," B.L. Turner, W.C. Clark, R.W.
Kates, J.F. Richards, J.T. Mathews and W.B. Meyer (eds.),
Cambridge University Press, Cambridge, U.K., p. 393-408.
Houghton, J.T., Jenkins, G.J. and J.J. Ephraums (1990)
Climate Change: The IPCC Scientific Assessment. Cambridge
University Press, Cambridge, 365 pp.
Houghton, J.T., L.G. Meira Filho, J. Bruce, L. Hoesung,
B.A. Callander, E. Haites, and K. Maskell (eds.) (1995) Climate
Change 1994: Radiative forcing of climate change and an
evaluation of the IPCC IS92 emission scenarios. Intergovernmental
Panel on Climate Change. Cambridge University Press, New York.
339 pp.
Jenkinson, D.S., Adams, D.E., and A. Wild
(1991) Model estimate of CO2 emissions from soil in response to
global warming. Nature, 351:304-306.
Jensen, M.E. and H.E. Haise (1963) Estimating
evapotranspiration from solar radiation. Journal of the
Irrigation and Drainage Division, 4:15-41.
Koner, C. (1993) CO2 fertilization: The
great uncertainty in future vegetation development. In:
"Vegetation Dynamics and Global Change," A.M. Solomon
and H.H. Shugart (eds.), Chapman & Hall Inc., New York, p.
53-70.
Kimball, B.A. (1975) Carbon dioxide and agricultural
yield: an assemblage and analysis of 430 prior observations. Agronomy
Journal, 75:779-788.
Leemans, R. and Cramer, W. P. (1990) The
IIASA climate database for land areas on a grid with 0.5 degree
resolution. WP-90-41, International Institute for Applied
Systems Analysis (IIASA), Laxenburg, Austria. 60 pp.
Manabe, S., and R.T. Wetherald (1987)
Large scale changes in soil wetness induced by an increase in
carbon dioxide. Journal of the Atmospheric Sciences, 44:1211-1235.
McGuire, A.D., Kicklighter, D.W., J.M. Melillo (1996a)
Global climate change and carbon cycling in grasslands and
conifer forests. In: "Global Change: Effects on Coniferous
Forests and Grasslands," J.M. Melillo and A.I. Breymeyer
(eds.), SCOPE volume (in press).
McGuire, A.D., J. M. Melillo, D.W. Kicklighter, Y. Pan, X.
Xiao, J. Helfrich, B. Moore III, C.J. Vorosmarty and A.L. Schloss
(1996b) The role of the nitrogen cycle in the global response of
net primary production and carbon storage to doubled CO2. Global
Biogeochemical Cycles (in review). McGuire, A.D., J.M.
Melillo, D.W. Kicklighter and L.A. Joyce (1995a) Equilibrium
response of soil organic carbon to climate change: Empirical and
process-based estimates. Journal of Biogeography, 22:785-796.
McGuire, A.D., J.M. Melillo and L.A. Joyce (1995b) The
role of nitrogen in the response of forest net primary production
to elevated atmospheric carbon dioxide. Annual Review of
Ecology and Systematics, 26: 473-503.
McGuire, A.D., L.A. Joyce, D.W. Kicklighter, J.M. Melillo,
G. Esser and C.J.Vorosmarty (1993) Productivity response of
climax temperate forests to elevated temperature and carbon
dioxide: A North America comparison between two global models. Climatic
Change, 24: 287-310.
McGuire, A.D., J.M. Melillo, L.A. Joyce, D.W. Kicklighter,
A.L. Grace, B. Moore III and C.J. Vorosmarty (1992) Interactions
between carbon and nitrogen dynamics in estimating net primary
productivity for potential vegetation in North America. Global
Biogeochemical Cycles, 6(2):101-124.
Melillo, J.M., I.C. Prentice, G.D. Farquhar, E.D. Schulze
and O.E. Sala (1996) Terrestrial biotic responses to
environmental change and feedbacks to climate. In: "Climate
Change 1995-The Science of Climate Change: Contribution of
Working Group I to the Second Assessment Report of the
Intergovernmental Panel on Climate Change," J.J. Houghton,
L.G. Meiro Filho, B.A. Callander, N. Harris, A. Kattenberg and K.
Maskell (eds.), Cambridge University Press, New York.
Melillo, J.M., D.W. Kicklighter, A.D. McGuire, W.T.
Peterjohn and K.M. Newkirk (1995) Global change and its effects
on soil organic carbon stocks. In: "Role of Nonliving
Organic Matter in the Earth's Carbon Cycle," R.G. Zepp and
Ch. Sonntag (eds.), John Wiley and Sons Ltd., p. 175-189.
Melillo, J.M. (1994) Modeling land-atmospheric
interaction: A short review. In: "Changes in Land Use and
Land Cover: A Global Perspective," W.B. Meyer and B.L.
Turner (eds.), Cambridge University Press, p. 387-409.
Melillo, J.M., A.D. McGuire, D.W. Kicklighter, B. Moore
III, C.J. Vorosmarty and A.L. Schloss (1993) Global climate
change and terrestrial net primary production. Nature, 363:234-240.
Melillo, J.M., T.V. Callaghan, F.I. Woodward, E. Salati
and S.K. Sinka (1990) Effects on ecosystems. In: "Climate
Change: The IPCC Scientific Assessment," J.T. Houghton, G.J.
Jenkins and J.J. Ephraums (eds.), Cambridge University Press, New
York, p. 283-310.
Mitchell, J.F.B., S. Manabe, T. Tokioka, V. and Meleshko
(1990) Equilibrium climate change and its implications for the
future. In: "Climate Change: The IPCC Scientific
Assessment," J.T. Houghton, G.J. Jenkins and J.J.Ephraums
(eds.), Cambridge University Press, New York, p. 131-164.
Mitchell, J.F.B., C.A. Senior and W.J. Ingram (1989) CO2
and climate: A missing feedback? Nature, 341:132-134.
Mooney, H.A., B.G. Drake, R.J. Luxmoore, W.C. Oechel and
L.F. Pitelka (1991) Predicting ecosystem responses to elevated
CO2 concentrations. BioScience, 2:96-104.
NCAR/NAVY (1984) Global 10-minute
elevation data. Digital tape available through National
Oceanic and Atmospheric Administration, National Geophysical Data
Center, Boulder.
Oechel, W.C. and G.H. Riechers (1986)
Impacts of increasing CO2 on natural vegetation, particularly
tundra. In: "Climate-Vegetation Interactions," C.
Rosenweig and R. Dickinson (eds.), OIES-UCAR Report OIES-2,
Boulder, CO.
Oechel, W.C. and G.H. Riechers (1987) Response of a tundra
ecosystem to elevated atmospheric carbon dioxide. U.S. Department
of Energy, Washington, D.C.
Pan, Y., A.D. McGuire, D.W. Kicklighter
and J.M. Melillo (1996) The importance of climate and soils for
estimates of net primary production: A sensitivity analysis with
the Terrestrial Ecosystem Model. Global Change Biology, 2:5-23.
Parton, W.J., J.M.O. Scurlock, D.S. Ojima, D.S. Schimel,
D.O. Hall and SCOPE GRAM group members (1995) Impact of climate
change on grassland production and soil carbon worldwide. Global
Change Biology, 1:13-22.
Post, W.M., W.R. Emanuel, P.J. Zinke and A.G.
Stangenberger (1982) Soil carbon pools and world life zones. Nature,
298:156-159.
Potter, C.S., J.T. Randerson, C.B. Field, P.A. Matson,
P.M. Vitousek, H.A. Mooney and S.A. Klooster (1993) Terrestrial
ecosystem production: a process model based on global satellite
and surface data. Global Biogeochemical Cycles,
7:811-841.
Prinn, R., H. Jacoby, A. Sokolov, C. Wang, X. Xiao, Z.
Yang, R. Eckaus, P. Stone, A. Ellerman, J. Melillo, J.
Fitzmaurice, D. Kicklighter, Y. Liu and G. Holian (1996)
Integrated global system model for climate policy analysis: I.
Model framework and sensitivity studies. Joint Program on the
Science and Policy of Global Change Report No. 7, Massachusetts
Institute of Technology, Cambridge, 76 pp.
Raich, J. W., E.B. Rastetter, J.M.
Melillo, D.W. Kicklighter, P.A. Steudler, B.J. Peterson, A.L.
Grace, B. Moore III and C.J. Vorosmarty (1991) Potential net
primary productivity in South America: Application of a global
model. Ecological Applications , 1(4):399-429.
Rosenzweig, C. and M.L. Parry (1994) Potential impact of
climate change on world food supply. Nature, 367(13):133-138.
Sarmiento, J.L. (1993) Atmospheric CO2
stalled. Nature, 365(21):697-698.
Schimel, D.S., B.H. Braswell, E.A. Holland, R. McKeown,
D.S. Ojima, T.H. Painter, W.J. Parton and A.R. Townsend (1994)
Climatic, edaphic and biotic controls over storage and turnover
of carbon in soils. Global Biogeochemical Cycles, 8(3):279-293.
Schimel, D.S. (1995) Terrestrial ecosystems and the carbon
cycle. Global Change Biology, 1: 1-15.
Schlesinger, W.H. (1977) Carbon balance in terrestrial
detritus. Annual Review of Ecology and Systematics, 8:51-81.
Siegenthaler, U. and J.L. Sarmiento (1993) Atmospheric
carbon dioxide and the ocean. Nature, 365(9):119-125.
Sokolov, A.P. and P.H. Stone (1995) Description and
validation of the MIT version of the GISS 2-D Model. Joint
Program on the Science and Policy of Global Change Report No. 2,
Massachusetts Institute of Technology, Cambridge, 46 pp.
Stone, P.H. and M.S. Yao (1987) Development of a
two-dimensional zonally averaged statistical-dynamical model.
Part II. The role of eddy momentum fluxes in the general
circulation and their parameterization. Journal of the
Atmospheric Science, 44(24):3769-3786.
Stone, P.H. and M.S. Yao (1990) Development of a
two-dimensional zonally averaged statistical-dynamical model.
Part III. The parameterization of the eddy fluxes of heat and
moisture. Journal of Climate, 3(7):726-740.
Townsend, A.R., P.M. Vitousek and E.A.
Holland (1992) Tropical soils could dominate the short-term
carbon cycle feedbacks to increased global temperature. Climatic
Change, 22: 293-303.
VEMAP Members (1995) Vegetation/Ecosystem
Modeling and Analysis Project (VEMAP): Comparing biogeography and
biogeochemistry models in a continental-scale study of
terrestrial ecosystem responses to climate change and CO2
doubling. Global Biogeochemistry Cycles, 9(4):407-437.
Ven Cleve, K., W.C. Oechel and J.L. Hom (1990) Response of
black spruce (Picea mariana) ecosystems to soil temperature
modification in interior Alaska. Canadian Journal of Forest
Research, 20:1530-1535.
Vorosmarty, C.J., B. Moore III, A.L. Grace, M.P. Gildea,
J.M. Melillo, B.J. Peterson, E.B. Rastetter and P.A. Steudler
(1989) Continental scale models of water balance and fluvial
transport: An application to South America. Global
Biogeochemical Cycles, 3(3): 241-265.
Wetherald, R.T. and S. Manabe (1988) Cloud
feedback processes in a general circulation model. Journal of
the Atmospheric Sciences, 45:1397-1415.
Willmott, C.J., M.R. Clinton and W.D. Philpot (1985)
Small-scale climate maps: a sensitivity analysis of some common
assumptions associated with grid-point interpolation and
contouring. The American Cartographer, 12:5-16.
Wilson, C.A. and J.F.B. Mitchell (1987) A doubled CO2
climate sensitivity experiment with a global climate model
including a simple ocean. Journal of Geophysical Research, 92(D11):
3,315-13,343.
Xiao, X., D.W. Kicklighter, J.M. Melillo,
A.D. McGuire, P.H. Stone and A.P. Sokolov (1996) Linking a global
terrestrial biogeochemical model and a 2-dimensional climate
model: Implications for the global carbon budget. Tellus (in
review).
Yao, M.S. and P.H. Stone (1987)
Development of a two-dimensional zonally averaged
statistical-dynamical model. Part I: The parameterization of
moist convection and its role in the general circulation. Journal
of the Atmospheric Sciences, 44(1):65-82.
Zhang, X. (1993) A vegetation-climate
classification system for global change studies in China. Quaternary
Sciences, 2:157-169. (in Chinese with English abstract)