
 |
The Iconology of a Humerus Tale by Farish Jenkins |
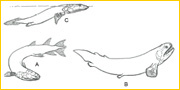 |
~A, B, C~ By Late Devonian times (~360 million years ago, mya), the rudimentary design of tetrapod limbs was established among sarcopterygian fishes that are known from diverse fossils from North America, Australia, eastern and western Europe. Only a few sarcopterygians survive today--lungfishes, with a species each in Africa, Australia and South America, and several species of the coelacanth Latimeria in the coastal waters of Africa and Indonesia. Although the humerus began as a simple element, flattened and block-like, a few features that are primitively present (an entepicondylar process, a ventral crest) persist throughout the evolutionary diversification of tetrapods. Thus the entepicondyle and deltopectoral crest of humans are piscine features. All sarcopterygians possessed bones that are homologous with radius and ulna. Few groups, however, had distal limb elements that are prototypic of digital rays (B, C). And only one group, the elpistostegids (C), not only had protodigits but also an array of cranial features antecedant to the tetrapod condition.1-7
|
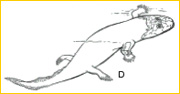 |
~D~ Acanthostega, retaining gills but with terrestrial limbs, is one of a number of primitive amphibians known from Late Devonian-Early Carboniferous deposits in Australia, eastern Europe, Greenland and North America. A flexed elbow supported the trunk, and the entepicondylar process enlarged to accommodate powerful antebrachial flexors. But the number of fingers remained distinctively fishy.8-9
|
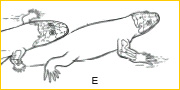 |
~E~ Humeral robustness and complexity increased as amphibians diversified in terrestrial habitats during the Late Paleozoic, and pentadactyly emerged as the tetrapod standard.10
|
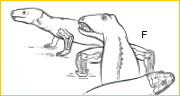 |
~F~ By the Late Carboniferous (~310 mya), the ancestral stocks of modern amniotes appeared, possessing in common the reproductive novelty of extraembryonic membranes– the amnion, chorion, and allantois. One primitive lineage, the sprawling pelycosaurs, evolved into both carnivorous and herbivorous niches. The humerus in predatory sphenacodontid pelycosaurs was of complex form, attesting to a differentiation of limb musculature and diversity of limb use.11
|
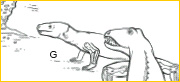 |
~G~ Therapsids, or mammal-like reptiles, descended from pelycosaurs and flourished during Permian and Triassic times (290-215 mya). Cynodonts known from South Africa and South America are evolutionarily the most advanced therapsids. In numerous dental, cranial, axial and appendicular features, cynodonts acquired anatomical patterns antecedent to the mammalian bauplan- including an upright, adducted limb posture and a prototypically mammalian humerus.12
|
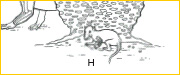 |
~H~ By the Late Triassic (~ 210 mya), true mammals appeared. Tiny in size, they remained for the most part diminutive throughout the remainder of the Mesozoic in a world dominated by dinosaurs and other reptiles.13
|
 |
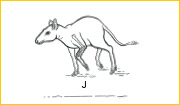
~I~ Substantial diversification of mammals began after the widespread extinctions at the close of the Cretaceous (65 mya). Among Paleocene mammals, condylarths represent some incipient specializations related to herbivory. Phenacodus possessed a bunodont dentition for browsing, and in place of claws, broad unguals functioned as proto-hooves. The robust skeletons of these heavy bodied, archaic forms reveal little of the cursorial adaptations developed by modern ungulates.14,15
|
 |
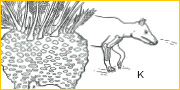
~J~ Among many groups of mammalian herbivores that have evolved, artiodactyls – or even-toed ungulates – are among the most successful in terms of taxonomic diversification. Diacodexis of the Early Eocene (55 mya) is the earliest known artiodactyl, close to the common ancestry of mouse deer, musk deer, deer, camels, pigs, hippos, giraffes, cows and antelopes. All artiodactyls share a distinctive, unique feature. The talus bears two trochleae; in addition to the conventional proximal trochlea of the tibio-talar joint, the talar head (articulating with the navicular) is also trochlear in form. This ‘double-pulleyed’ talus is part of a spring mechanism that contributes to the prodigious running and bounding abilities of many artiodactyls.16
|
 |
~K~ Elomeryx, an anthracotheriid artiodactyl from the Late Eocene through Late Oligocene of Europe and North America (38–23 mya), is placed in the superfamily Hippopotamoidea, but possesses premolars that are suggestively similar in form to those of the earliest known whales.17,18
|
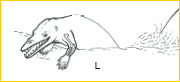 |
~L~ Along the shores of the once vast Tethys Sea (of which the present day Mediterranean is but a remnant), primitive whales appeared during the Eocene. First recognized from fragmentary skulls, recent finds in Pakistan have turned up vertebrae and limbs. The hindfeet of these amphibious archaeocetes had a talus with two trochleae -- whales are derived artiodactyls!19-21
|
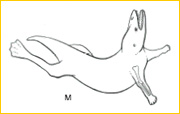 |
~M~ Rhodocetus, a primitive archaeocete cetacean found in Eocene deposits of Pakistan, swam by paddling with powerful hind limbs.21
|
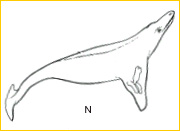 |
~N~ Durodon, an advanced basilosaurid cetacean known from Late Eocene sediments in Egypt, abandoned hind limbs in favor of flexing and extending the caudal fluke, a mechanism employed by all modern whales.22
|
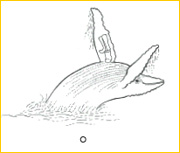 |
~O~ A humpback whale, Megaptera novaeangliae (‘the big wing of New England’), breaches with an exuberant back flip in Cape Cod coastal waters. As with so many other mammals, whales are social and communicative, but have otherwise transformed every other aspect of their mammalian heritage into an elegant, perfected return to the sea. In some, teeth have been abandoned in favor of keratinous sheets of baleen, sieving plankton from 45,000-liter gulps of water or benthic life from 2-20 m2 mouthfuls of mud and sand sucked from the oceanic floor. Yet the humerus has reverted to a simple structural form, as in their sarcopterygian ancestors of the Devonian.
|
| ~References~
|
|