| Figure 2
Nature Cell Biology
1, 493 - 499 (1999)
Published online: 8 November 1999; | doi:10.1038/70281
Cooperative symmetry-breaking by actin polymerization in a model for
cell motilityAlexander van Oudenaarden
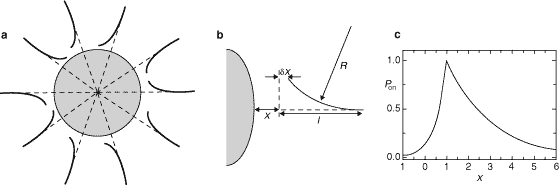
& Julie A. Theriot | | | | Figure 2. Description of the multifilament stochastic model.
a, Initial symmetric distribution of actin filaments around a bead.
The filaments are assumed to be orientated perpendicular to the bead surface
and are evenly spaced. b, Each actin filament is considered as an elastic
rod with length l. The tip−bead distance for a non-bent filament
is given by x. Because of thermal fluctuations, the end of an actin
filament that is close to the bead surface can bend, which increases the bead−tip
distance by ‘Ąx. The other end of the filament is fixed in space
because of crosslinking with other filaments. For small deflections, the filament
bends with a constant radius of curvature R. c, The probability
of subunit addition, P
on, is a function of distance x. Note that all distances are
normalized to the size of the gap‘î required to permit intercalation of an actin
monomer between the filament tip and the bead surface (‘î’âŕ 2.7 nm)3.
A necessary condition for subunit addition to the free end is that the gap opened
between the fluctuating tip and the bead surface be large enough to allow a monomer
to intercalate: x + ‘Ąx ’†1.
For x <1, the
probability that this condition be met can be calculated from Boltzmann's
distribution for thermal bending. Experimentally, actin polymerization has
been seen to be confined to the immediate vicinity of the bacterial or bead
surface14,
16. This is represented by introducing a exponential
decay of P
on(x) for x >1. P
on(x) is maximal at x = 1, where the filament tip is
at the position closest to the bead that allows actin-monomer intercalation without
filament bending. See Methods for details.
|
| | |  | | |
 |
|  |
|