A Brief Introduction to Bacteriophage P22
Assembly
Barrie Greene
Viruses have long been known as sources of human disease, but have
now also become important tools of biological research used in DNA
cloning, targeted gene therapy, and phage display of novel proteins.
Although viruses are extremely diverse in their life cycles and
infectious mechanisms, most share a common structure, consisting of
an inner core of condensed nucleic acid enclosed within a spherical
protein capsid. The structures of these capsids are based upon
icosahedral symmetry, as illustrated in these images
of viruses.
In the case of herpesviruses and adenoviruses, as well as the
double-stranded DNA bacteriophages such as P22, the initial product
of the viral assembly pathway is not an infectious virion but a
closed shell that does not contain DNA. These precursor shells, or
procapsids, include proteins not found in the mature virion, but
essential for their production. These proteins are termed
"scaffolding proteins".
During the 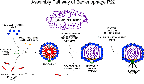 assembly pathway of bacteriophage P22, 300 molecules of a 33 kD
scaffolding protein coassemble with the 420 coat protein subunits to
form a double-shelled structure with the scaffolding inside. The
procapsid also includes a dodecameric portal complex at one vertex,
which serves as the channel through which the DNA enters. In
addition, there are from 10-20 molecules each of three pilot
proteins, needed for injection of DNA into the host cell.
assembly pathway of bacteriophage P22, 300 molecules of a 33 kD
scaffolding protein coassemble with the 420 coat protein subunits to
form a double-shelled structure with the scaffolding inside. The
procapsid also includes a dodecameric portal complex at one vertex,
which serves as the channel through which the DNA enters. In
addition, there are from 10-20 molecules each of three pilot
proteins, needed for injection of DNA into the host cell.
Upon the commencement of DNA packaging, all 300 scaffolding molecules
are released intact from within the shell, presumably through the
channels present at the fivefold and sixfold centers. The DNA is
pumped into the capsid in a process that involves two virally-encoded
proteins and ATP. During these events the capsid undergoes
conformational changes that result in expansion, angularization, and
closure of the channels. See the P22 procapsid
structure at 19 ™ resolution  .
.
The arrangement of the scaffolding subunits within the procapsid is
not known; it may not match the icosahedral symmetry of the coat
shell. No structures of scaffolding proteins from any virus have been
solved.
The Role of Scaffolding Protein in Phage P22 Capsid Assembly
and Maturation
My work focuses on both understanding the roles played by the
scaffolding subunits in the complex processes of virus assembly and
maturation, and identifying regions of the molecule required for
specific functions.
Scaffolding release can be reproduced in vitro in the absence of DNA
by low concentrations of guanidine hydrochloride. This release from
within the capsid was reversible, permitting study of scaffolding
interactions with the assembled coat lattice. The rapidity of
scaffolding reentry suggested that the subunits could rebind to
specific sites within the capsid. Procapsids contained two classes of
scaffolding subunits, which may represent binding to different sites
within the lattice; thus, all scaffolding proteins do not make
equivalent interactions with the coat protein. These sites became
lost or inaccessible upon capsid expansion and maturation.
Image reconstruction from cryo-electron micrographs of procapsids
containing scaffolding suggest that although the scaffolding subunits
interact with the capsid lattice, the scaffolding core is not
arranged with the same icosahedral symmetry as the outer shell. This
work was done in collaboration with Drs. Chiu and Prasad and their
colleagues at Baylor College of Medicine.
Pam Thuman-Commike , a graduate student at Baylor, has a page
which discusses image reconstructions of phage P22 structures in more
detail.
A set of new missense mutants at four sites in the scaffolding
protein were isolated and characterized. Their locations are shown on
a map of the bacteriophage P22
scaffolding sequence 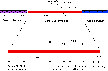 . These mutants did not prevent capsid assembly under
restrictive conditions. Two mutants were defective in incorporation
of the portal complex which serves as the channel through which DNA
is packaged. These mutations may identify a region of the protein
required for interaction with the portal. Two mutants in a different
region of the sequence were impaired in scaffolding release both in
vivo and in vitro. These mutations may identify a new domain required
for scaffolding release. Both these mutations resulted in severe
destabilization of part of the protein to thermal denaturation.
Scaffolding release appeared to be required for capsid expansion; in
turn, scaffolding release seemed to depend upon the presence of a
portal. This may help to order the pathway of events in phage
maturation.
. These mutants did not prevent capsid assembly under
restrictive conditions. Two mutants were defective in incorporation
of the portal complex which serves as the channel through which DNA
is packaged. These mutations may identify a region of the protein
required for interaction with the portal. Two mutants in a different
region of the sequence were impaired in scaffolding release both in
vivo and in vitro. These mutations may identify a new domain required
for scaffolding release. Both these mutations resulted in severe
destabilization of part of the protein to thermal denaturation.
Scaffolding release appeared to be required for capsid expansion; in
turn, scaffolding release seemed to depend upon the presence of a
portal. This may help to order the pathway of events in phage
maturation.
A scaffolding region required for binding to coat protein was
identified by functional assays of proteolytic fragments, and found
to be the C-terminal 20-30 residues. The structural organization of
the purified wild-type and mutant proteins were also probed by
protein denaturation techniques. The unfolding of scaffolding protein
was not a simple two-state mechanism as for a typical globular
protein, but a complex process invoving the sequential denaturation
of multiple domains. The mutant amino acid substitutions selectively
destabilized particular domains, allowing them to be assigned to
known functions. The scaffolding protein was notably unstable, to the
extent that some domains are probably largely unstructured at
physiological temperatures. The less stable domains include the
regions involved in coat binding, portal insertion and scaffolding
release, suggesting that many critical scaffolding functions may
require a high degree of conformational flexibility.
Based on these results I propose a model for assembly in which a
terminal region of the scaffolding protein induces conformational
changes in the coat protein leading to efficient polymerization. The
release of scaffolding may not be a passive result of changes within
the coat lattice but involve active conformational changes within the
scaffolding subunits in response to signals associated with DNA
packaging. Docking of the DNA packaging complex at the portal could
cause a signal to be propagated throughout the scaffolding core
resulting in scaffolding release, after which the coat lattice is
freed to expand to its mature conformation.
Related Publications:
Greene, B., and King,
J. (1994). Binding
of scaffolding subunits within the P22 procapsid lattice.
Virology 205, 188-197.
Greene, B. and King,
J. (1996). Scaffolding
mutants identifying domains required for P22 procapsid assembly and
maturation. Submitted to Virology.
Thuman-Commike, P. A, Greene, B., Jokana, J., Prasad, B. V. V.,
King, J., Prevelige, P.
E., and Chiu, W. (1996). Three-dimensional
structure of scaffolding-containing phage P22 procapsids by electron
cryo-microscopy. Submitted to J. Mol.
Biol.
Return to King
Lab Research
Return to King Lab
Homepage