Imaging and Elasticity of the Stalk of the Protozoan Peritrich Vorticellid :
An Ultrafast Biological Supramolecular Spring
Rafael E. Bras
(*Funded by the Dupont-MIT Alliance in conjunction with
Profs. Alexander van Oudenaarden (PHYSICS-MIT), Paul T. Matsudaira
(Biology/BEH/Whitehead Institute-MIT), and L. Mahadevan
(MECHE-MIT/PHYSICS-CAMBRIDGE U.))
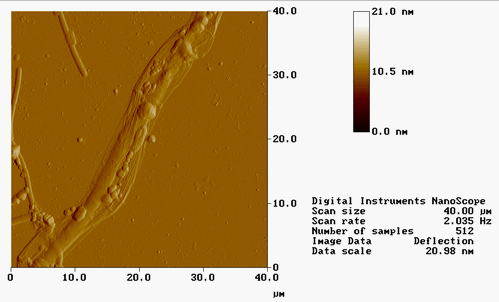
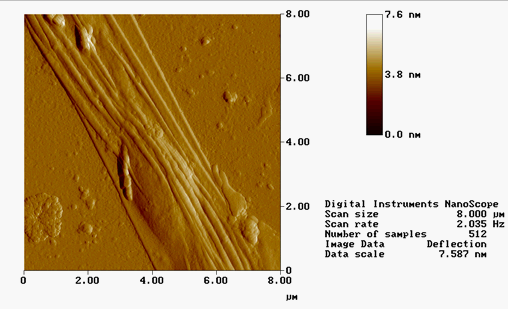
AFM IMAGES FROM FALL 2001 :
(*Contact mode in air on mica)
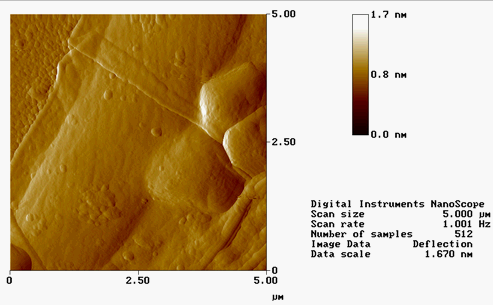
AFM IMAGES FROM SUMMER 2001 :
(*Contact mode in air on polylysine-coated mica)
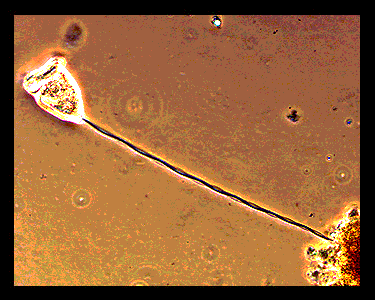
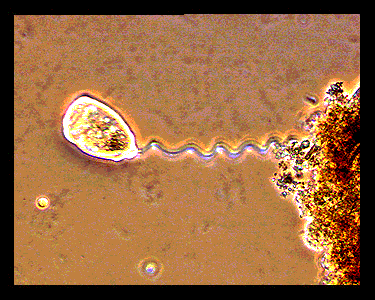
(*Optical Micrographs BELOW obtained from : http://bioweb.uwlax.edu/zoolab/Table_of_Contents/Lab-02/Vorticella/Live_Vorticella_1/live_vorticella_1.htm)
Nature has found various ingenious ways to induce movement on the cellular and
molecular scale. One of the most powerful and fastest engines is found in
stalked peritrichs of the family Vorticellids (Vorticella, Carchesium,
Zoothamnium). These organisms are composed of a zooid (cell body, diameter
~50 microns) and a long stalk (in its extended state 20-200 microns long).
One end of the stalk is connected to the zooid and the other end attaches to
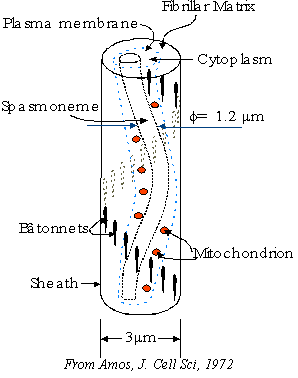
a solid support such as a submerged leaf. The spasmoneme is the
organelle responsible for the remarkable motility and lies within the stalk.
After addition of Ca2+ the spasmoneme rapidly contracts at velocities
approaching 8 cm/s (Amos, 1971; Moriyama et al., 1998).
The contraction is assumed to be a response effective in protecting
the cell against dangerous mechanical disturbances such as turbulence
in the surrounding water. This rate of contraction is extremely high,
the zooid traverse ~1800 times its length per second. This is 120 times
larger than a bacterial flagellar engine and 360 times larger than a
midsize car engine at 60 mph. The spasmoneme is located
asymmetrically with the stalk and stalk sheath is
stiffened on the opposite side by complex extracellular fibers (Amos, 1972).
Due to this asymmetry the stalk bends rather than collapses when the
spasmoneme contracts.
Birefringence measurements (Amos, 1971) and electron microscopy
(Amos, 1972; Allen, 1973; Amos, 1975) indicate that spasmoneme in
the presence of Ca2+ (fully extended state) consists of a bundle of
roughly parallel filaments (Fig. 2B) that are weakly crosslinked to each other.
In the absence of calcium ions, the extended spasmoneme collapses as an entropic spring. An early model (Hoffman-Berling, 1958) assumes that energy is stored by electrostatic repulsion between negatively charged filaments. By adding Ca2+ the negative charges are screened, which results in a collapse into the isotropic state. The structure and chemical composition of the spasmoneme are unknown. Recent experiments indicate the importance a major spasmoneme protein, called spasmin, that potentially binds Ca2+ ions and orchestrates the contraction (Asai et al., 1998; Maciejewski et al., 1999). Although the Vorticella system has been explored for more than three centuries now, starting with the observations of the 17th-century microscopist Leewenhoek (van Leeuwenhoek, 1676), the basic biochemistry and biophysics of this powerful spring remains unresolved. The overall goals of this project is to elucidate the molecular mechanism of spasmoneme contraction by (i) biochemical characterization of the constituents of the spasmoneme; (ii) biophysical characterization of the mechanics
and dynamics of the stalk; and (iii) development of theoretical models
to quantitatively understand the contraction mechanism.
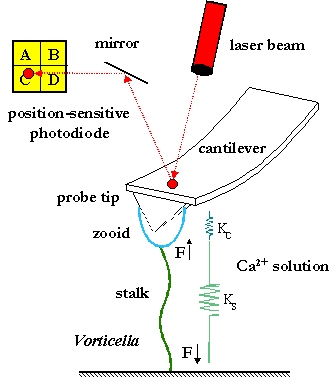
In order to test theoretical predictions of stalk contraction quantitative
biomechanical experiments are underway which involve measuring the
elastic properties (force-extension curves) of individual Vorticella stalks
(Asai et al., 1978) at different Ca2+ concentrations using high-resolution
force spectroscopy. The experimental force-extension curves will be fitted
to known theoretical models. Relevant models are rubber-like elasticity models
(Treloar, 1975; Moriyama et al., 1999), entropic strings models such as the
worm-like chain model (Marko et al., 1995), and novel theoretical models.
The worm-like chain model is particularly successful in describing
collapsed DNA molecules in the presence of multivalent cations
(Baumann et al., 2000). Possible conformational changes of protein
subunits in the stalk will significantly alter the force-extension curves.
REFERENCES:
Allen, R. D. Structures linking the myonemes, endoplasmic reticulum, and surface membranes in the contractile ciliate Vorticella. J. Cell Biol. 56, 559-579 (1973).
Amos, W.B. Reversible mechanomechinal cycle in the contraction of Vorticella. Nature 229, 127-128 (1971).
Amos, W. B. Structure and coiling of the stalk in the peritrich ciliates Vorticella and Carchesium. J. Cell Sci. 10, 95-122 (1972).
Amos, W. B. Contraction and calcium binding in the Vorticellid ciliates. Molecules and Cell Movement. S. Inoue, and R. E. Stephens, editors. Raven Press, New York. 411-436 (1975).
Asai, H., T. Ochiai, K. Fukui, M. Watanabe, and F. Kano. Improved preparation and cooperative calcium contraction of glycerinated Vorticella. J. Biochem. 83, 795-798 (1978).
Asai, H., T. Ninomiya, R. Kono, and Y. Moriyama. Spasmin and a putative spasmin binding protein(s) isolated from solubilized spasmonemes. J. Eukaryot. Microbiol. 45, 33-38 (1998).
Baumann, C. G., V. A. Bloomfeld, S. B. Smith, C. Bustamante, M.D. Wang, and S. M. Block. Stretching of single collapsed DNA molecules. Biophys. J. 78, 1965-1978 (2000).
Hoffmann-Berling, H. Der Mechanismus eines neuen, von der Muskelkontraktion verschiedenen Kontraktionszyklus. Biochim. Biophys. Acta. 27, 247-255. (1958).
Jones, A. R., T. L. Jahn, and J. R. Fonseca. Contraction of protoplasm. IV. Cinematographic analysis of the contraction of some petririchs. J. Cell Physiol. 75, 9-20 (1970).
Katoh, K., M. Kikuyama. An all-or-nothing rise in cytosolic Ca2+ in Vorticella Sp. J. Exp. Biology 200, 35-40 (1997).
Khoklov et al; Adv. Poly. Sci. 109, 123 (1993).
Landau, L., and E. Lifshitz, The Theory of Elasticity, Butterworth-Heinemann, Boston (1995).
Maciejewski, J.J., E.J. Vacchiano, S.M. McCutcheon, H.E. Buhse Jr. Cloning and expression of a cDNA encoding a Vorticella convallaria spasmin: an EF-hand calcium-binding protein. J. Eukaryot. Microbiol. 46, 165-173 (1999).
Mahadevan, L. and P. Matsudaira. Motility powered by supramolecular springs and ratchets. Science 288, 95-99 (2000).
Marko, J. F. and E. D. Siggia. Stretching DNA. Macromolecules 28, 8759-8770 (1995).
Moriyama, Y., S. Hiyama, and H. Asai. High-speed video cinematographic demonstration of stalk and zooid contraction of Vorticella convallaria. Biophys. J. 74, 487-491 (1998).
Moriyama, Y., H. Okamoto, and H. Asai. Rubber-like elasticity and volume changes in the isolated spasmoneme of giant Zoothamnium sp. under Ca2+-induced contraction. Biophys. J. 76, 993-1000 (1999).
Treloar, L. R. The physics of rubber elasticity, 3rd Ed. Clarendon Press, Oxford (1975).
van Leeuwhoek, A. Philos. Trans. R Soc. London Ser. B 12, 133 (1676).