Workshop Proceedings: Short Papers
Application of Passive Acoustic Methods for Detection, Location and Tracking of Whales
Christopher W. Clark
Bioacoustics Research Program, Cornell Lab of Ornithology, 159 Sapsucker Woods Rd., Ithaca, NY 14850 cwc2@cornell .edu, http://www.birds.cornell.edu/brp/
Introduction
Baleen whales produce species specific sounds. At least five species produce long, patterned, hierarchically organized sequences of sounds referred to as songs. Baleen whale distribution and relative abundance estimates are traditionally based on visual surveys from vessels and/or airplanes. This approach is limited by visibility conditions and access to observation platforms. Acoustic monitoring using either single or multiple sensors offers a significant improvement by increasing spatial and temporal sampling. Throughout the last 20 years, acoustic hardware and software tools have been developed and applied to survey whale species and gain insights into natural behaviors. This includes the use of Navy SOSUS arrays to detect and estimate numbers of vocal animals throughout an ocean basin, autonomous seafloor sensors to detect, locate and track species of interest in regions, and sparse or towed hydrophone arrays to detect, locate and track selected species in specific study areas. For at least four species, SOSUS data reveal annual seasonal and large geographic fluctuations reflecting migration, feeding and breeding patterns. For bowheads combined visual-acoustic efforts have lead to calculation of a robust population assessment and trend over a 20-year period, as well as understandings of acoustic functions. For blue and fin whales, integrated approaches combining passive acoustic methods with visual, biopsy, photo-ID and prey field surveys are beginning to reveal critical details of behavioral ecology and significant insights into vocal functions.
Background
The Cornell Bioacoustics Research Program specializes in the development and application of advanced techniques to investigate the mechanisms and evolutionary bases of animal acoustic behaviors. Techniques successfully developed for the study of one organism or taxonomic group are often applied to another group. Part of the motivation for including the topic of whales in this symposium was to provide an opportunity to exchange information with researchers primarily interested in fishes. Clearly many of the tools developed for studies on whales are applicable to fish. Species in both taxa produce rich assortments of sounds in the low-frequency band (<1000Hz). Most sounds are species specific and can be used as indicators of species presence and relative abundance. Males are known to produce acoustic displays associated with breeding. Similar sound propagation models can be applied to quantify probabilities of detection. An integrated approach in which acoustic techniques are combined with other methods (e.g., visual, molecular, oceanographic) can yield tremendous advancements in our collective understandings of marine biology.
Whale bioacoustics
All 11 species of baleen whales produce sounds. Representations of sound repertoires are complete for the five coastal species: bowhead, gray, humpback and the southern and northern right whales. There are good representations for two pelagic species, blue and fin whales. Whale source levels (RMS power in dB re 1µPa @ 1 m) have been reported as high as 188 dB (tabulated in Richardson et al. 1995). Under certain conditions (e.g., water depth that is not well matched to the signal’s frequency band of lowest transmission loss, or highly reverberant environments), there is little or no advantage to increased source level, and selection should favor changes in other acoustic features to optimize communication effectiveness and range.
In the marine environment, assuming a selective advantage for long-range communication, the influences of physical acoustics should have imposed strong selective pressures on the acoustic features of communication sounds. There is evidence in support of this general hypothesis on sound function when considering the physical acoustic properties of the ocean environment in combination with the acoustic features of sounds produced by mysticete whales. Two primary acoustic properties that strongly influence communication range are transmission loss (TL) and ambient noise. In a shallow water habitat (< 200-300m) sounds in the 100-800Hz band experience the lowest TL and there is often a window of low ambient noise in this frequency range. Along a shelf break or in deep ocean environments (> 1000m), sounds below 100Hz experience the lowest TL and between 10-50Hz there is a plateau of low ambient noise. The acoustic features for species which breed and forage in predominantly coastal habitats are very different from those from a pelagic environment, as illustrated in Figure 1.
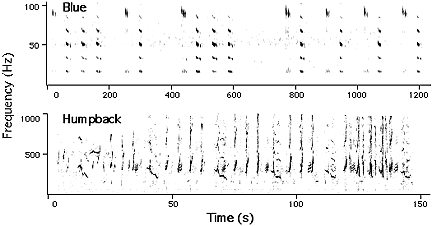 Figure 1. Spectrograms of blue and humpback songs to illustrate differences in acoustic characteristics for species predominantly from pelagic (blue) and coastal (humpback) habitats. |
Ambient noise levels are different for these two environments, and the dominant frequency band of a species’ song is generally matched to the frequency band of low ambient noise as shown in Figure 2 (Clark and Ellison in press).
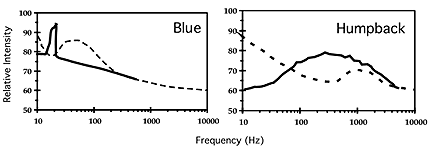 Figure 2. Spectra (solid lines) for pelagic (blue) and coastal (humpback) species overlaid on ambient noise spectra (dashed lines) for the two different habitats. |
Traditional visual survey methods are inadequate for documenting species presence/absence, distribution or relative abundance. The spatial and temporal sampling scales required are prohibitive for ship or aerial surveys, but passive acoustic sampling can offer an effective solution. Over the last ten years the acoustic activity of four species have been monitored on an ocean-scale basis in the north Pacific and North Atlantic using the Navy’s SOSUS network (Watkins et al.2000, Charif et al. 2001). Results provide large scale patterns of vocal activity throughout the year in areas where animals (e.g., humpbacks) are known to breed. Surprisingly, however, for pelagic species with no known breeding or calving grounds, singing occurs throughout most of the year even during the feeding season in high latitude areas (Figure 3).
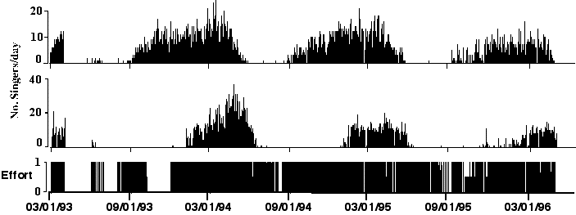 Figure 3. Daily counts of blue (top panel) and humpback (bottom panel) singers detected on SOSUS arrays in the western North Atlantic over a three year period. |
For regional-scale sampling, autonomous seafloor recorders have been used in conjunction with visual surveys (aircraft or vessels) to estimate species densities and with active acoustic surveys to study the relationship between whales and the physical and biological oceanographic conditions. The results from such integrated efforts are immensely more informative than any one survey method alone. The number of cases in which acoustics detected species first and more often than visual survey is increasing, even for species that are plentiful and highly visible.
In cases where a highly focused effort is required, for example, when conducting a visual-acoustic census or testing specific hypotheses regarding sound function, a variety of hydrophone systems can be used. This includes sparse arrays of hydrophones, distributed networks of autonomous seafloor recorders or a towed beamforming arrays (Clark et al. 1996; Clark and Fristrup 1997; Croll et al. 2002). Such techniques provide mechanisms to continuously describe the number, locations and movements of individual animals in great detail. When combined with data from other focused field methods such as photo-id and biopsy sampling, one can relate age, sex, breeding status and behavior of individuals within a population to their acoustic behaviors (e.g., Croll et al. 2002). Integrated approaches offer tremendous opportunities for expanding critical knowledge in such diverse areas as marine vertebrate mating strategies and human impacts on the marine environment.
Acknowledgements
The basis for this synthesis comes from several decades of research sponsored by the National Geographic Society, New York Zoological Society, National Science Foundation, Naval Research Lab, North Slope Borough Department of Wildlife Management, and the Office of Naval Research. Jack Bradbury, Donald Croll, William Ellison, Leila Hatch and Roger Payne provided stimulating discussions. I am indebted to all the staff in the Bioacoustics Research Program for assistance with data collection, analysis, and interpretation.
References
Charif, R. A., Clapham, P. J. and Clark, C.W. 2001. Acoustic detections of singing humpback whales in deep waters off the British Isles. Mar. Mamm. Science 17(4):751-768.
Clark, C. W., Charif, R., Mitchell, S., and Colby, J. 1996. Distribution and Behavior of the Bowhead Whale, Balaena mysticetus, Based on Analysis of Acoustic Data Collected During the 1993 Spring Migration off Point Barrow, Alaska. Rep. int. Whal. Commn. 46: 541-552.
Clark, C. W., and Ellison, W. T. In press. Potential use of low-frequency sounds by baleen whales for probing the environment: evidence from models and empirical measurements. in Echolocation in Bats and Dolphins (J. Thomas, C. Moss and M. Vater, eds.). The University of Chicago Press.
Clark, C.W. and Fristrup, K. 1997. Whales ‘95: A combined visual and acoustic survey of blue and fin whales off southern California. Rep. Int. Whal. Commn. 47:583-600.
Croll, D. A., Clark, C. W., Acevedo, A., Tershy, B., Flores, S., Gedamke, J. and Urban, J. 2002. Only males fin whales sing loud songs. Nature 417:809.
Richardson, W. J., C. R. Greene, Jr., C. I. Malme, and D. H. Thomson. 1995. Marine Mammals and Noise. Academic Press, New York, 576 pp.
Watkins, W. A., M. A. Daher, G. M. Reppucci, J. E. George, D. L. Martin, N. A. DiMarzio, and D. P. Gannon. 2000. Seasonality and distribution of whale calls in the North Pacific. Oceanography 13:62-67.s