|
Using a Towed Array to Survey Red Drum Spawning Sites in the Gulf
of Mexico
Scott A Holt
University of Texas at Austin Marine Science Institute
750 Channel View Drive, Port Aransas TX 78373 USA.
sholt@utmsi.utexas.edu
Introduction
The red drum (Sciaenops ocellatus) is an important recreational
and, in some locations, commercial species throughout its range. Juveniles
generally live in estuaries and move to nearshore oceanic waters as
they reach maturity (Pearson 1929). Adults range widely over the nearshore
continental shelf waters throughout the year but apparently move to
coastal waters to spawn (Overstreet 1983). Spawning is generally thought
to take place in coastal waters near inlets (Jannke 1971, Holt et al.
1985) although Lyczkowski-Shultz et al. (1988) found eggs and larvae
out to 34 km from shore in the eastern Gulf of Mexico. There is also
evidence of limited spawning activity within estuaries in Florida (Murphy
and Taylor 1990, Johnson and Funicelli 1991) and in North Carolina (Luczkovich
et al. 1999).
The location of spawning areas has typically been inferred through capture
of fish with mature gonads or the distribution of eggs and larvae. Red
drum make loud, characteristic sounds during spawning (Guest and Lasswell
1978). Listening for the characteristic sound production has recently
been used to locate red drum spawning sites in Indian River Lagoon,
Florida (Johnson and Funicelli 1991), and in Plamico Sound, North Carolina
(Luczkovich et al. 1999), and at tidal inlets in South Carolina (personal
communication, M. R. Collins, South Carolina Department of Natural Resources).
These surveys have been done with both hand-held hydrophones and remotely
placed sonobuoys.
Over a four-year period from 1998-2001, a hydrophone mounted on a pier
in the Aransas Pass, Texas tidal inlet has been use to record sounds
of red drum spawning activity every evening during the September through
October spawning period. Recordings were made for 20 s every 15 m from
1700 to 0100 hours and spanned the 4-5 hour evening spawning period
of red drum (Holt et al. 1985). Red drum produced characteristic spawning
sounds from about one hour before sunset to about three hours after
sunset with the most intense activity occurring during the two hours
following sunset (S. Holt, unpublished data). These data, along with
collections of red drum eggs and larvae at the site, confirmed that
red drum spawn actively in the vicinity of the tidal inlet. The spatial
extent of red drum spawning was still unknown but it was clear that
surveying sound production during spawning was an effective means of
locating spawning sites.
This paper reports on a survey of potential spawning sites in the nearshore
western Gulf of Mexico using a towed hydrophone array.
Study Area and Methods
The survey was conducted in the northwestern Gulf of Mexico along the
central portion of the Texas, USA, coast. Preliminary surveys with a
hand-held hydrophone in the area revealed that red drum spawning sounds
were more commonly observed along the 10 m contour than in either shallow
water near the surf zone or farther offshore in deeper water. Hence,
for this initial survey, three transects were established roughly parallel
to the coastline along the 10 m contour. Transects were sampled on three
consecutive nights (one transect per night) in late September 2000.
Sampling commenced about 30 - 45 min before sunset, which was about
1925, and ran for about 3.5 hours.
The towed array was composed of eight hydrophones in an 80 meter cable
connected to a 200 meter towing cable and was towed at approximately
4.5 kts from a 105 foot stern trawler. The array is spectrally flat
(i.e. no peaks in sensitivity) from 6Hz to 18 kHz, with a sensitivity
of approximately -191 dB re 1 volt per m
Pa at 7.2 kHz. The signals from each of the eight separate hydrophones
were saved to an eight-track digital recorder (Tascam DA-88) sampling
at 44 kHz. The combination of a temporal window of spawning vocalizations
(about 3.5 hours) and optimum towing speed for the array of (4.5 kts)
limited each nightly transect to about 20 km.
Red drum produce low frequency sounds described as knocks (Fish and
Mowbray 1970) or drumming (Guest and Lasswell 1978). Although Guest
and Lasswell (1978) found the "dominant energy" of their recordings
from a tank was around 240 Hz — 1000 Hz, I have found the fundamental
frequency of red drum calls obtained from unconstrained fish in the
field to consistently be around 140 Hz — 160 Hz (Fig. 1).
 |
| Figure 1.
Sonogram of a red drum call from an unconstrained individual in
the field. This particular call consists of three widely spaced
knocks followed by two pairs of closely spaced knocks. |
Each call consists of a variable number of pulses, or
knocks, that are repeated at a range of pulse repetition rates (Guest
and Lasswell 1978, laboratory observations; S Holt unpublished data,
field observations). Whether there are specific behaviors associated
with specific call types is unknown but the existence of numerous variants
in call pattern suggests individual variability. Despite variation in
call duration and pulse repetition rate, the consistency in fundamental
frequency and general character of the call pattern make recognition
by ear relatively easy.
Recorded signals from the array were analyzed by listening to the tapes
while observing the real-time power spectra and real-time sonogram on
a computer screen (SpectraPro 3.32, Sound Technology Inc.). Two classes
of red drum sounds could be distinguished. One was a low frequency rumble
with a prominent energy peak in the 150 Hz range. This was presumed
to be from large numbers of red drum producing sounds simultaneously
but at some distance from the hydrophone. (The sound produced by the
ship and the hydrophone itself was determined to have dominant energy
in the range of 250 Hz — 300 Hz.) The other class of sounds was
clearly distinguishable calls made by an individual or small group of
red drum.
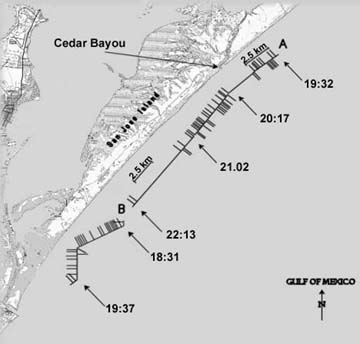 |
| Figure 2. Location San Jose "A" and "B"
hydrophone transects. The line indicates the cruise track. Bars
above the line indicate low one-minute drumming rates at that location.
Bars below the line indicate high one-minute drumming rates. Sampling
time is indicated randomly along the track. |
The occurrence of background rumble indicates spawning
activity in the vicinity of the hydrophone but more work is needed before
the spatial scale over which those sounds travel can be meaningfully
interpreted. For this paper, I will describe only the distribution of
individual or small-group calls. From our observations and the work
of Luczkovich et al. (1999), it appears that the drumming of an individual
red drum can be distinguished over a distance of about 100 m. Thus,
we can roughly define the spatial distribution of individual red drum
detected by the hydrophones as a 200 m swath along the transect. The
physical location of each observation was determined by comparing the
underway data recorded from the ship’s SAIL system (which included
time and latitude/longitude as well as several physical parameters)
and the clock time on the digital recorder which was carefully synchronized
with the ships clock. The data set was initially constructed by recording
the hour/minute/second of each identifiable call. The data was then
summarized by counting the number of calls heard in each one-minute
segment (the ships location was recorded once per minute so that was
our finest scale of spatial resolution). The number of calls/minute
was arbitrarily divided in two groups: <16 per minute and 16 or more
per minute. This division was set to separate the typically lower occurrence
of drumming (5-10 per minute was typical) from the relatively rarer
higher rate (we rarely heard more than 20-30 per minute). Finally the
drumming rate (i.e. none, low, or high) was plotted on the cruise track.
Results
Red drum calls were detected along most sections of the three transects
(Figs. 2 & 3). Calls were detected both in extensive clusters and
in isolated occurrences along the transects. For example, on the San
Jose "A" transect (Fig. 3), there are two occurrences of near
continuous calling that extend over several kilometers. On the same
transect, there are several isolated occurrences of red drum calls and
extensive segments (up to 4 km) where there are no calls. Transect segments
were dominated by the absence of red drum calls. There was a total of
474 minutes of observations over all transects. Of those, 330 minutes
(70%) had no red drum calls, 109 minutes (23%) had low drumming rates
(<16 per min), and 35 minutes (7%) had high drumming rates
(>15 per min). High drumming activity was concentrated
in two segments along the San Jose "A" transect and in one
segment of the Matagorda transect. One segment, on the east end of the
transect, spanned 5 minutes of towing time and covered 600 m. The other,
farther to the west on that transect, spanned 14 minutes of towing time
and covered 2.2 km. Only 4 of the 14 minutes in this segment were low
level drumming and none were without drumming.
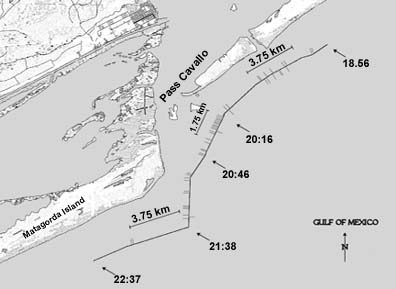 |
| Figure 3. Location of the Matagorda
hydrophone transect. See Fig.2 legend for details. |
The most intense drumming activity occurred between 1830 and 2130. Little
drumming was heard after 2130 on the Matagorda or San Jose "A"
transects (data for the later part of the San Jose "B" transect
was lost due to an audio tape malfunction). Low and high drumming rates
were distributed throughout this time period without any temporal pattern.
Discussion
Based on the distribution of sound production, red drum appear to spawn
all along the nearshore region of the central Texas coast. This survey
was not spatially comprehensive enough to fully delineate the spawning
area, but it is clear from this initial survey that spawning activity
is widespread. Spawning was not concentrated at inlets as suggested
by earlier authors (Simmons and Breuer (1962), Jannke (1971). Areas
of the coastline far removed from the inlets had relatively intense
drumming activity and confirms suggestions of Murphy and Taylor (1990)
that spawning also occurs over the nearshore continental shelf.
It is still not exactly clear how drumming by male red drum should be
interpreted. There are at least three possibilities: 1) the drumming
male will engage in spawning at that location on that evening; 2) the
drumming male is calling from a potential spawning site but will spawn
at that site on that day only if joined (or selected) by a cooperative
female; or 3) the drumming male may move to another place before engaging
in spawning. Luczkovich et al. (1999) observed instances of red drum
drumming without finding eggs and Johnson and Funicelli (1991) found
red drum eggs without hearing drumming. In both instances, short-term
observations were made in shallow water with a hand held hydrophone
and the observers may have disturbed the fish or missed part of the
spawning process. At this point, it is assumed that drumming roughly
equates to spawning but the issue needs more investigation.
The distribution of drumming male red drum suggest that some, if not
most, of the spawning takes place among widely distributed individuals
as opposed to highly aggregated groups. Only 7% of the one-minute summaries
recorded high drumming rates of more than 15 calls per minute. Guest
and Lasswell (1978) reported a call rate of about 2-16 calls per minute
for captive red drum in courtship. Our subjective impression from listening
to the tapes was that many of the low drumming rates were produced by
a single fish. There were, however, at least two large aggregations
of drumming fish. Both were in the vicinity of Cedar Bayou, a relatively
small but historically persistent tidal inlet. One of these aggregations
spanned a linear distance of over 2 km and its breath was undetermined.
The number of calls per minute (up to 40) indicates that several red
drum were calling simultaneously within the roughly 100 meter detection
range of the hydrophones and this "density" was consistent
over most of the 2 km stretch.
The full extent of the offshore spawning area of red drum is yet to
be determined and much remains to be learned about their reproductive
strategies, but the use of towed hydrophone arrays offers promise of
an efficient means to achieve those goals.
Acknowledgments
I thank John Keller for processing the audio data and Cameron Pratt
for preparing the figures. The crew of the R/V Longhorn was instrumental
in acquiring the recordings. This work was funded through a grant from
the Sid W. Richardson Foundation.
References
Fish, M. P., and W. H. Mowbray. 1970. Sounds of western North Atlantic
fishes; a reference file of biological underwater sounds. Johns Hopkins
Press, Baltimore.
Guest, W. C., and J. L. Lasswell. 1978. A note on courtship behavior
and sound production of red drum. Copeia 1978: 337-338.
Holt, G. J., S. A. Holt, and C. R. Arnold. 1985. Diel periodicity of
spawning in sciaenids. Marine Ecology Progress Series 27: 1-7.
Jannke, T. E. 1971. Abundance of young sciaenid fishes in Everglades
National Park, Florida, in relation to season and other variables. University
of Miami Sea Grant Technical Bulletin 11: 128p.
Johnson, D. R., and N. A. Funicelli. 1991. Spawning of the red drum
in Mosquito Lagoon, East-Central Florida. Estuaries 14: 74-79.
Luczkovich, J. J., H. J. Daniel, III, and M. W. Sprague. 1999. Characterization
of critical spawning habitats of weakfish, spotted seatrout and red
drum in Pamlico Sound using hydrophone surveys. Pages 128. North Carolina
Department of Environment and Natural Resources, Morehead City, NC.
Lyczkowski-Shultz, J., J. P. Steen, Jr., and B. H. Comyns. 1988. Early
life history of red drum (Sciaenops ocellatus) in the northcentral Gulf
of Mexico. Pages 148. Mississippi-Alabama Sea Grant Consortium.
Murphy, M. D., and R. G. Taylor. 1990. Reproduction, growth, and mortality
of red drum Sciaenops ocellatus in Florida waters. Fishery Bulletin,
US 88: 531-542.
Overstreet, R. M. 1983. Aspects of the biology of the red drum, Sciaenops
ocellatus, in Mississippi. Gulf Research Reports Supplement 1: 45-68.
Simmons, E. G., and J. P. Breuer. 1962. A study of redfish, Sciaenops
ocellata Linnaeus and black drum, Pogonias cromis Linnaeus. Publications
of the Institute of Marine Science 8: 184-211.
Return
to Top | Workshop Proceedings: Short
Papers
|