|
IS ACOUSTIC
CALLS A PREMATING REPRODUCTIVE BARRIER BETWEEN TWO NORTH-EAST ATLANTIC
COD (Gadus morhua) GROUPS – A REVIEW
Jarle Tryti Nordeide* and Jens Loss Finstad+
* Faculty of Fisheries and Natural Sciences, Bodø Regional University,
N-8049 Bodø, Norway. + Master student at: Department of Fisheries
and Marine Biology, University of Bergen, Norway
Summary
This paper reviews the first attempts to test the hypothesis that spawning
calls of male migratory "Arctic" and a stationary "Coastal"
cod group, which are sympatric during the spawning season, is a premating
behavioural reproductive barrier. As predicted from the hypothesis,
a hushed hubbub of sound with a transient character, band-width and
harmonic spacing typical of cod calls, was revealed at a major spawning
ground during the spawning period but not six months later. Moreover,
individual calls from male cod kept in tanks varied a lot both in harmonic
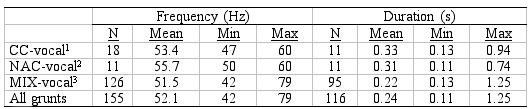
spacing and duration, from 42 to 79 Hz and 0.11 to 1.25 s, respectively.
Such individual variation in calls is expected if females choose mate
on the basis of their calls. However, the results so far has failed
to support the third prediction, since no differences have been found
between the calls of the two groups, neither in harmonic spacing, duration,
or temporal structure of the calls.
Introduction
North-east Atlantic cod consist of two stocks, the North-east Arctic,
or "Arctic" cod, and the Norwegian coastal cod, or "Coastal"
cod (Rollefsen 1934). Arctic cod migrates from the feeding areas in
the Barents Sea to the spawning areas along the Norwegian coast and
the most important spawning area is off the Lofoten Islands where the
main spawning occurs in March and April (Bergstad & al. 1987). Coastal
cod inhabit coastal areas and fjords, migrates short distances and spawns
in a large number of fjords along the Norwegian coast (Rollefsen 1954),
including off the Lofoten Islands (Hylen 1964). Both cod groups spawn
in March and April and mature speciemens are sympatric during spawning
at the major spawning grounds off the Lofoten Islands (Nordeide 1998).
A controversial topic during decades has been whether or not the two
cod groups interbreed, and a majority of studies conclude that they
rarely do (see references in Nordeide & Pettersen 1998). If so,
active partner choice is required, and lekking has recently been suggested
to best describe the cod’s mating system (Hutchings & al. 1999;
Nordeide & Folstad 2000).
Møller (1968) suggested that active partner choice based on acoustic
calls may be an behavioural mechanism which prevents interbreeding between
the two cod groups. The aim of this paper is to summarize the first
attempts to test predictions derived from Møller’s hypothesis.
The predictions are that (i) recordings from major cod spawning grounds
should reveal sound with characteristics typical of cod whereas much
less sound should be revealed outside the spawning season, (ii) calls
from individual cod should show considerable variation, and (iii) calls
from Arctic and Coastal cod should differ.
Material and methods
To study sound at a major spawning ground, recordings were carried out
during the night at five stations off the Lofoten Islands at 68ê13.0’N
14ê38.7’E in Northern Norway, during the spawning season 8. and
9. April 1997, and half a year later on 4. September 1997 (Nordeide
& Kjellsby 1999). The measuring hydrophone with a 32 dB gain built-in
preamplifier had a total sensitivity of –152 dB re 1 V/mPa within
the frequency range of 16 Hz to 2 kHz. In order to emphasize the transient
character of the sound the digital recordings were analysed with Short-time
Fourier Techniques (STFT), at the Norwegian Defence Research Establishment
(Nordeide & Kjellsby 1999).
To compare the calls from the two groups, recordings were carried out
in land-based tanks in 1998 to 2001 (Finstad, in prep.). Speciemens
of the Arctic and Coastal cod were caught by trawl and transferred to
tanks where recordings were carried out during the spawning period in
1998 - 2001. The smallest male used was 53 cm and the largest male was
94 cm long, whereas the smallest and largest females were 51 cm and
104 cm long, respectively. The average length of the five cod groups
varied from 74.2 cm to 84.0 cm for males, and from 68.8 cm to 90.5 cm
for females. Most recordings were from three 6 m diameter fibreglass
tanks, but a 3 m diameter fibreglass tank was also used. Water level
was 1.4 – 1.5 m in all tanks. Recording equipment was a 1 inch
piezoceramic spherical hydrophone with a sensitivity of –198 dB
ref 1V 1mPa, a Levell preamplifier type TA 601 with 60 dB gain, and
a Sony TCD-D100 digital tape recorder. In 1998, the recordings were
carried out with 12 speciemens (six Coastal males) in the experimental
tank, whereas seven Arctic cod (5 males) where present in 1999. After
the first years of experience we had identified two major problems:
(i) we were not able to identify which cod produced the calls, and (ii)
relatively few grunts had been recorded. In 2000 and 2001 we therefore
chose to first record grunts with all cod in each group kept together,
to increase the number of grunts. Thereafter, we split the groups of
fish into a total of 10 smaller sub-groups to increase the minimum number
of individual cod which could possibly produce the grunts. The groups
in 2000 and 2001 consisted of 8 (3 males), 25 (19 males) and 22 (7 males)
cod, respectively. The sub-groups consisted of from 4 (2 males) to 16
(11 males) cod. Towards the end of the spawning season the fish were
killed. Examination of their otoliths by the Institute of Marine Research
in Bergen, revealed that the recordings were carried out with groups
and sub-groups consisting of (i) only male Coastal cod with or without
the presence of Arctic females, (ii) only male Arctic cod with or without
the presence of Coastal females, and (iii) a mixture of Coastal and
Arctic males and females. These three alternative combinations are referred
to as "Coastal-vocal", "Arctic-vocal" and "Mix-vocal"
groups respectively, since only males produce sound during the spawning
period (Brawn 1961c, Hawkins & Rasmussen 1978, see also Templeman
& Hodder 1958, Engen & Folstad 1999). The number of individual
cod (statistical "N") which could have produced the recorded
grunts were minimum 3 and maximum 12 in the "Coastal-vocal"
group, minimum 2 and maximum 7 in the "Arctic-vocal", and
minimum 4 and maximum 22 in the "Mix-vocal" groups. Analysis
by Avisoft-SASLabPro v. 3.74 provided estimates of harmonic spacing
and duration of the grunts. Temporal structure of 78 recorded high quality
grunts were analysed from oscillograms. Parameters included were number
of downward peaks, time-intervals between peaks, and duration of the
grunt. These parameters were analysed by Principal Component Analysis
by the software "The Unscrambler" v. 7.5.
Results and Discussion
Field recordings provide support for the hypothesis that acoustic
communication is important during cod spawning. Sound recordings at
the major spawning ground off the Lofoten Islands revealed a hushed
hubbub of sound, at approximately 40 – 500 Hz during the spawning
period (Fig. 1a). Much less sound was revealed in September (Fig. 1b)
when no cod spawn and migratory cod had emigrated to the Barents Sea.
Nordeide & Kjellsby (1999) argues that this sound most likely is
made by spawning cod since (1) The sound activity is highest in the
frequency range where it has been suggested cod communicate (Chapman
& Hawkins 1973; Hawkins & Rasmussen 1978). (2) The sound above
50 Hz had a transient character as expected for cod grunts. (3) More
than ca. 50 million male cod spawned off the Lofoten Islands in April
1997, and recordings were made where the Institute of Marine Research
in Bergen located the highest densities of spawning cod. (4) Cod totally
dominated, by constituting more than 98 % by wet weight, the experimental
and commercial catches in the area the seven days before, during and
after the recordings (Nordeide & Kjellsby 1999).
Variation in calls between individual cod kept in tanks is as expected
under the hypothesis that acoustic communication is important during
female mate choice. Grunts from cod kept in tanks vary in harmonic spacing
from 42 Hz to 79 Hz, and in duration from 0.11 s to 1.25 (Table 1).
The two grunts with the lowest and highest harmonic spacing came from
two different individuals, since they were recorded from two different
sub-groups. The shortest and longest calls were also produced by two
different individuals. This shows that different cod individuals may
grunt at different frequencies and durations, but we cannot tell anything
about each individual cod’s possibility to vary their calls.
The average harmonic spacing of grunts from the Coastal-vocal groups
and Arctic-vocal groups were 53.4 Hz and 55.7 Hz, respectively, and
the calls from the two cod groups lasted on average 0.33 s and 0.31
s, respectively (Table 1). The difference of 2.3 Hz in harmonic spacing
and 0.02 s in duration between the two groups, is probably negligible.
The difference cannot be tested statistically because the grunts are
not independent events, since we were not able to tell which cod produced
each call. Moreover, the calls from the Mix-vocal groups show no bimodal
distribution in harmonic spacing or duration, as is expected if Coastal
and Arctic cod call at two separate frequencies or durations. In the
multivariate analysis of the temporal structure of the grunts, the first
and second principal component explained 45 and 20%, respectively, of
the total variation. However, the analysis did not cluster the grunts
from Arctic-vocal and Coastal-vocal cod into two separate groups, as
should be expected if the temporal structure of the two cod-groups differed
(Finstad, in prep.). The hypothesis thus failed to pass the third test,
since we have not been able to separate the calls from the two cod groups.
However, analysis of temporal structure will continue with less rough
analytical tools.
References
Bergstad, O.A., T. Jørgensen, O. Dragesund 1987. Life history
and ecology of the gadoid resources of the Barents Sea. – Fisheries
Research 5:119-161.
Brawn, V.M. 1961c. Sound production by the cod (Gadus callarias L.).-
Behaviour 18:239-255.
Chapman, C.J. & A.D. Hawkins 1973. A field study of hearing in the
cod, Gadus morhua L. – Journal of Comparative Physiology. 85: 147-167.
Engen, F. & I. Folstad 1999. Cod courtship song: a song at the expense
of dance?- Canadian Journal of Zoology 77:542-550.
Finstad, J.L. In prep. Acoustic calls of Norwegian coastal cod and North-east
Arctic cod (Gadus morhua). Cand. scient. thesis at the Departure of
Fisheries and Marine Biology, University of Bergen. (Planned to be delivered
in May 2002).
Hawkins, A.D. & K.J. Rasmussen 1978. The calls of Gadoid fish. –
Journal of the Marine Biology Association of U.K. 58:891-911.
Hutchings, J.A., T.D. Bishop & C.R. McGregor-Shaw 1999. Spawning
behaviour of Atlantic cod, Gadus morhua: evidence of mate competition
and mate choice in broadcast spawner.- Canadian Journal of Fisheries
and Aquatic Sciences 56:97-104.
Hylen, A. 1964. Coastal cod and skrei in the Lofoten area. - Fiskeridirektoratets
Skrifter Serie Havundersøkelser, 13:27-42.
Møller, D. 1968. Genetic diversity in spawning cod along the
Norwegian coast.- Hereditas 60:1-32.
Nordeide, J.T. 1998. Coastal cod and north-east Arctic cod – do
they mingle at the spawning grounds in Lofoten?- Sarsia 83:373-379.
Nordeide, J.T. & I. Folstad 2000. Is cod lekking or a promiscuous
group spawner?- Fish and Fisheries 1:90-93.
Nordeide, J.T. & E. Kjellsby 1999. Sound from spawning cod at their
spawning grounds.- ICES Journal of Marine Science 56:326-332.
Nordeide, J.T. & I.H. Pettersen 1998. Haemoglobin frequencies and
vertebrae numbers of cod (Gadus morhua L.) off northern Norway –
test of a population structure hypotheses.- ICES Journal of Marine Science
55:134-140.
Rollefsen, G. 1934. The cod otolith as a guide to race, sexual development
and mortality.- Rapports et Proces-Verbaux des Reunions du Conseil International
Pour l’Exploration de la Mer 88:1-6.
Rollefsen, G. 1954. Observations on the cod and cod fisheries of Lofoten.-
Rapports et Proces-Verbaux des Reunions du Conseil International Pour
l’Exploration de la Mer, 136:40-47.
Templeman, W. & V.M. Hodder 1958. Variation with fish length, sex,
stage of sexual maturity, and season in the appearance and volume of
the drumming muscles of the swimbladder in the haddock, Melanogrammus
aeglefinus (L.).- Journal of the Fisheries Research Board of Canada.
15:355-390.
Table 1
Mean, minimum and maximum values of frequency and duration of grunts
from CC-vocal1, NAC-vocal2 and the MIX-vocal3 group. Number of grunts
(N) of frequency measurements and duration measurements differ because
some of the duration measurements were difficult to perform due to background
noise. The table is from Finstad (in prep.).
 |
|
1
: Grunts from groups and sub-groups consisting of CC males and
CC and NAC females, based on otolith analyses
2 : Grunts from groups and sub-groups consisting of
only NAC individuals, based on otolith analyses
3 : Grunts from groups and sub-groups consisting of
both CC and NAC, both males and females, based on otolith analyses
|
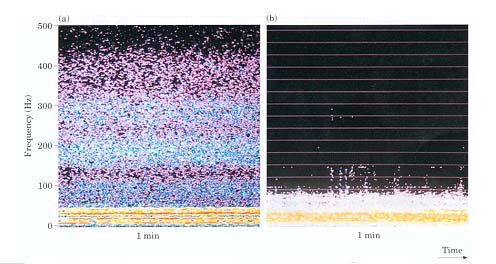 |
| Figure 1.
Recordings from a major spawning ground off the Lofoten Islands,
(a) during the spawning period in April and (b) in September when
no cod spawn. Reprinted from ICES Journal of Marine Science,Vol.
56, Nordeide, J.T. & E. Kjellsby, Sound from spawning cod at
their spawning grounds, 326-332, 1999, by permission of the publisher
Academic Press. |
Return
to Top | Workshop Proceedings: Short
Papers
|